-
O copiar el link


Filtros de Contribuciones
Buscar Contribución:
por Mauricio Montero Astúa, Izayana Sandoval Carvajal, Lisela Moreira Carmona, William Villalobos Muller, Laura Garita Salazar, Sofía Carvajal Rojas
Recibido: 30/7/2023 – Publicado: 30/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-3
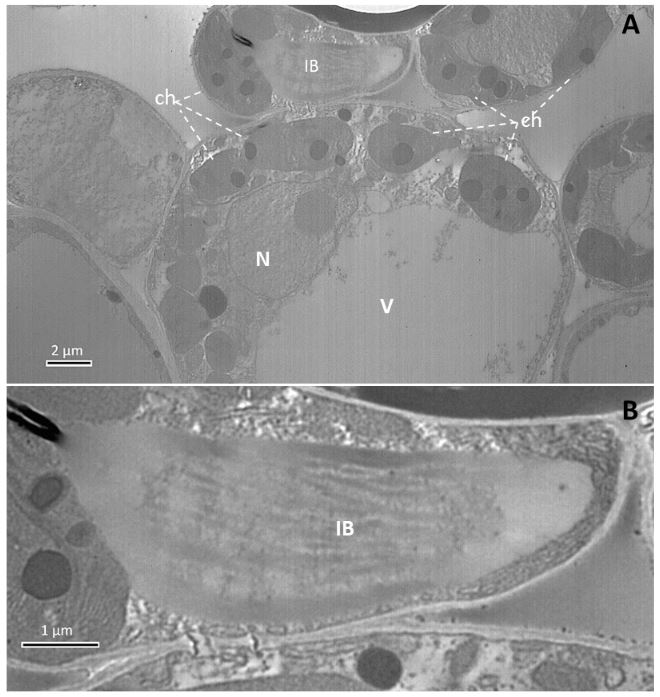
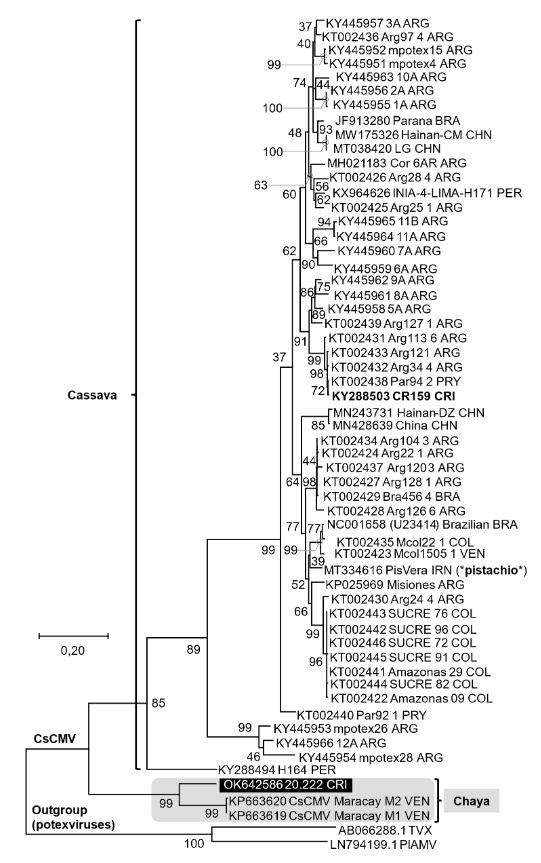
Resumen Antecedentes/Objetivo. Las hojas del arbusto de chaya (Cnidoscolus aconitifolius) o árbol espinaca, o chicasquil (en Costa Rica), son parte de la tradición culinaria Mesoamericana, con origen en el sur de México y Guatemala. El objetivo de este trabajo fue verificar la naturaleza viral de la enfermedad de una planta de chaya con mosaico detectada e identificar la especie del virus.
Materiales y Métodos. La detección viral se realizó mediante TEM, RT-PCR y secuenciación parcial empleando imprimadores degenerados para potexvirus. Se realizaron pruebas de patogenicidad mediante inoculaciones mecánicas empleando plantas de Nicotiana benthamiana y de chaya.
Resultados. Se reporta la detección del CsCMV en una planta de chaya con síntomas de mosaico. La patogenicidad y asociación del virus con los síntomas se demostraron mediante su inoculación en Nicotiana benthamiana y en plantas de chaya. Nuestra hipótesis es que corresponde a una introducción reciente del virus y se discute cómo las tradiciones culturales influyen en la distribución de los virus de plantas.
Conclusión. Los hallazgos confirman la presencia de un virus relacionado al CsCMV, previamente no informado para Costa Rica, en Cnidoscolus aconitifolius. En este trabajo resaltan la necesidad de estudiar su distribución y diversidad a través de Latinoamérica
-
O copiar el link

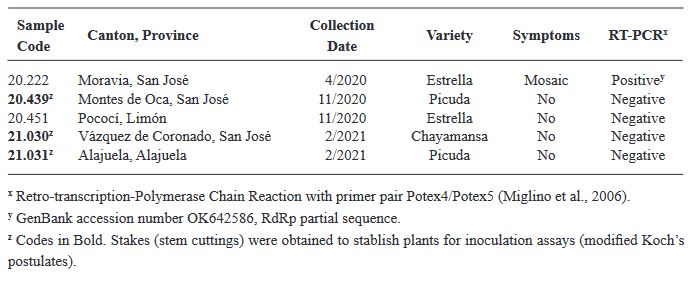
por Erika J. Zamora Macorra, Katia Aviña Padilla, Rosemarie W Hammond, Daniel L. Ochoa Martínez
Recibido: 23/8/2023 – Publicado: 23/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-5
Resumen Antecedentes/Objetivo. El virus del fruto rugoso marrón del jitomate (ToBRFV) ha surgido como una amenaza significativa para los cultivos de la familia Solanaceae, incluidos el tomate y el pimiento. Su presencia en México desde 2018 ha generado preocupación sobre su impacto en la producción agrícola. La detección temprana y precisa de este patógeno es crucial para prevenir su propagación y mitigar sus efectos. En México, se emplean varias técnicas moleculares para su diagnóstico, incluyendo RT-PCR convencional, RT-qPCR y RT-qPCR multiplex.
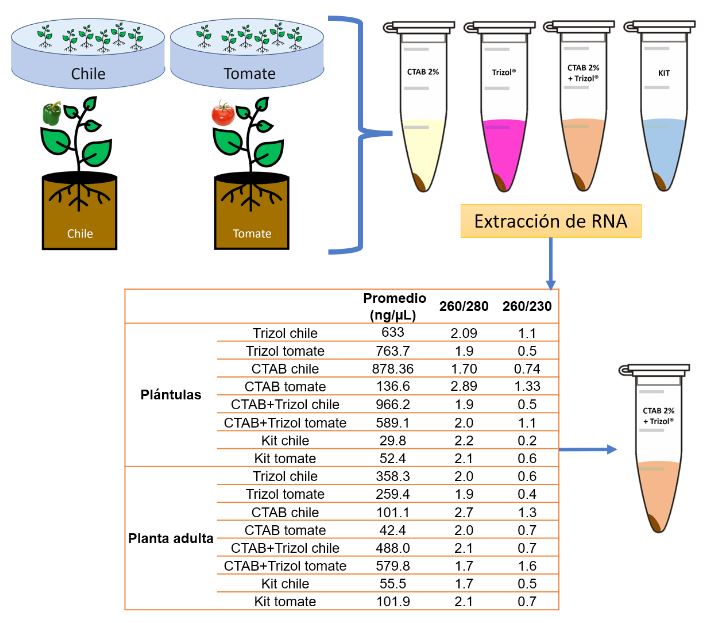
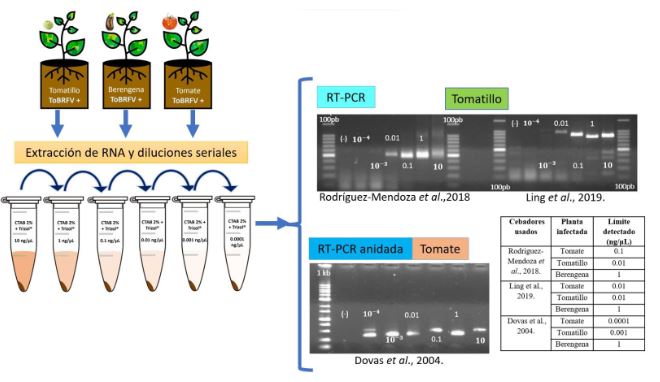
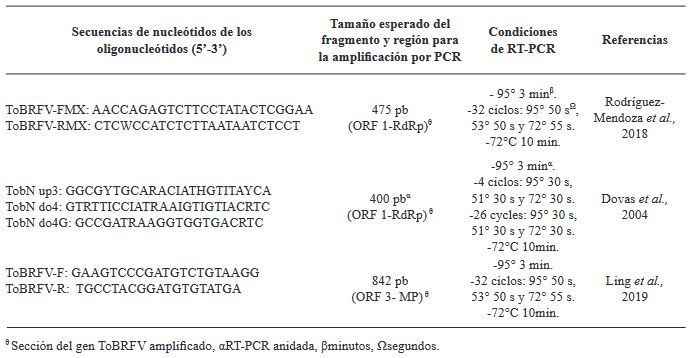
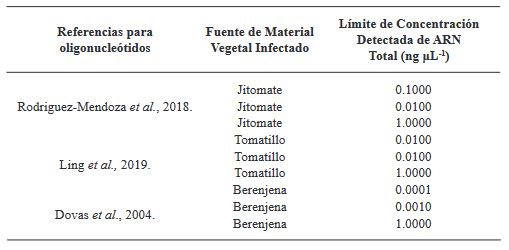
Materiales y Métodos: El objetivo de esta investigación fue evaluar la eficiencia de diferentes métodos de extracción de ARN en combinación con oligonucléotidos PCR específicos para la detección de ToBRFV.
Resultados. Entre los métodos probados, el protocolo de extracción de ARN CTAB-Trizol combinado con PCR anidada utilizando oligonucleótidos reportados por Dovas et al. (2004) se identificó como el método molecular más sensible para detectar el virus.
Conclusión. Este hallazgo destaca la importancia de seleccionar la combinación adecuada de protocolos de extracción y amplificación para lograr la sensibilidad y precisión óptimas en la detección de ToBRFV. Palabras clave: Virus del fruto rugoso marrón del jitomate, ToBRFV, cultivos de Solanaceae, extracción de ARN, RT-PCR, detección molecular, producción agrícola.
-
O copiar el link

por Carlos D. Ramos Villanueva, Guadalupe Carrillo Benitez, Erika J. Zamora Macorra, Eduardo Santiago Elena, Samuel Ramírez Alarcón, Jezrael Jimenez Vidals, Ricardo Ricardo López
Recibido: 31/7/2023 – Publicado: 19/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-1
Resumen Antecedentes y objetivo: El Tomato brown rugose fruit virus (ToBRFV) es uno de los principales patógenos del cultivo de jitomate en México. A pesar de los esfuerzos para evitarlo, es casi imposible por el bajo porcentaje de transmisión que tiene en semilla y la gran facilidad para ser transmitido mediante las labores culturales; por lo tanto, se buscan alternativas de manejo. Esta investigación tuvo como objetivo determinar el efecto de microorganismos endófitos aplicados al suelo, en plantas de jitomate infectadas por el ToBRFV.
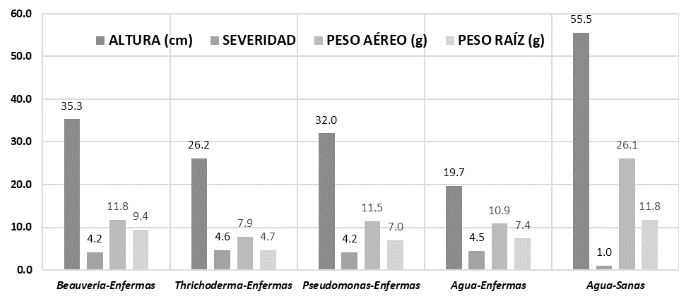
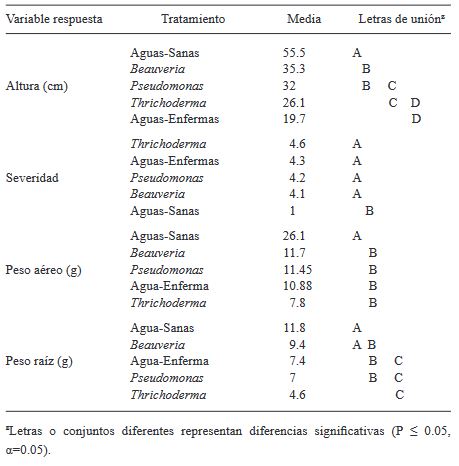
Materiales y Métodos. Se utilizó una planta de jitomate como unidad experimental, con 13 repeticiones por tratamiento. Los tratamientos en plantas de jitomate infectadas con ToBRFV fueron Beauveria peruviencis, Trichoderma longibrachiatum, Pseudomonas sp. y agua como testigo enfermo; también se incluyó un tratamiento de plantas sanas tratadas con agua como testigo absoluto. Las variables respuesta fueron altura de la planta, peso fresco de la parte aérea y de la raíz y severidad (dos evaluaciones). Las mediciones se analizaron mediante pruebas HSD de Tukey-Kramer por cada par.
Resultados. Se encontraron diferencias significativas entre tratamientos: Beauveria peruviencis, Trichoderma longibrachiatum, Pseudomonas sp. y agua como testigo enfermo. El tratamiento que favoreció el desarrollo de las plantas infectadas (79 % más altas y 15 % con más peso que el testigo infectado) y disminuyó su severidad fue B. peruviencis, seguido de Pseudomonas sp. El tratamiento que provocó menor desarrollo de la planta (31% menos que el testigo infectado) e inclusive aumentó la severidad fue T. longichrachiatum
-
O copiar el link

por Erika Janet Zamora Macorra, Norma Ávila Alistac, Erika Lagunes Fortiz, Sergio de los Santos Villalobos
Recibido: 30/8/2023 – Publicado: 28/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-7
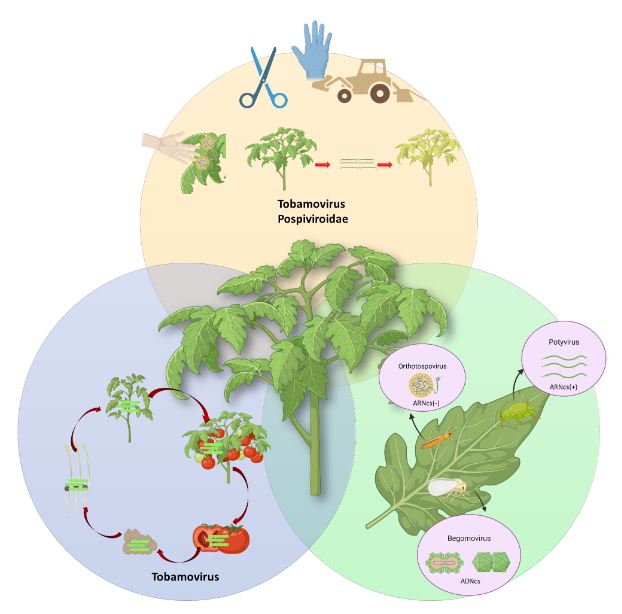
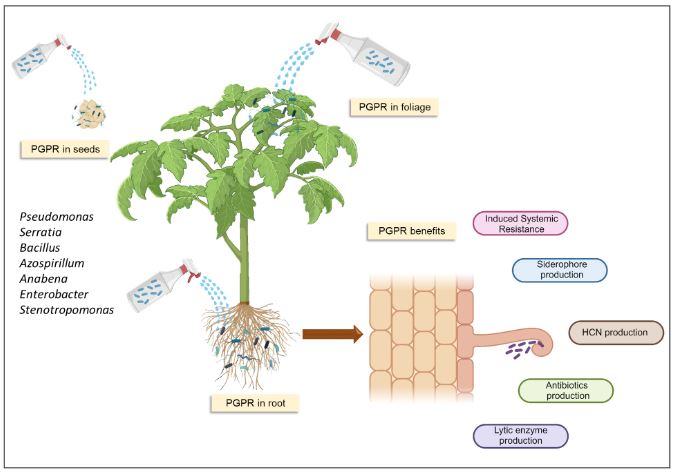
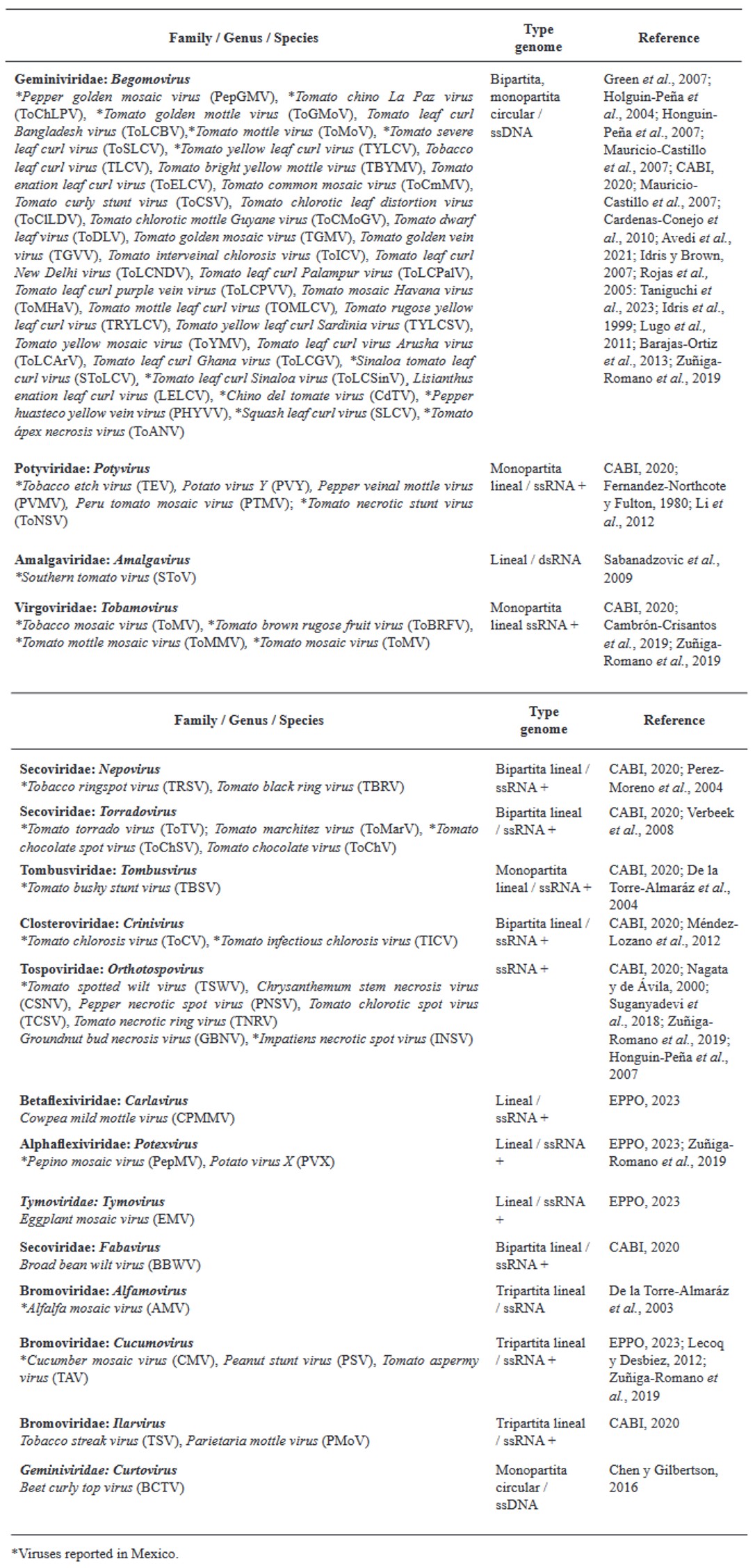
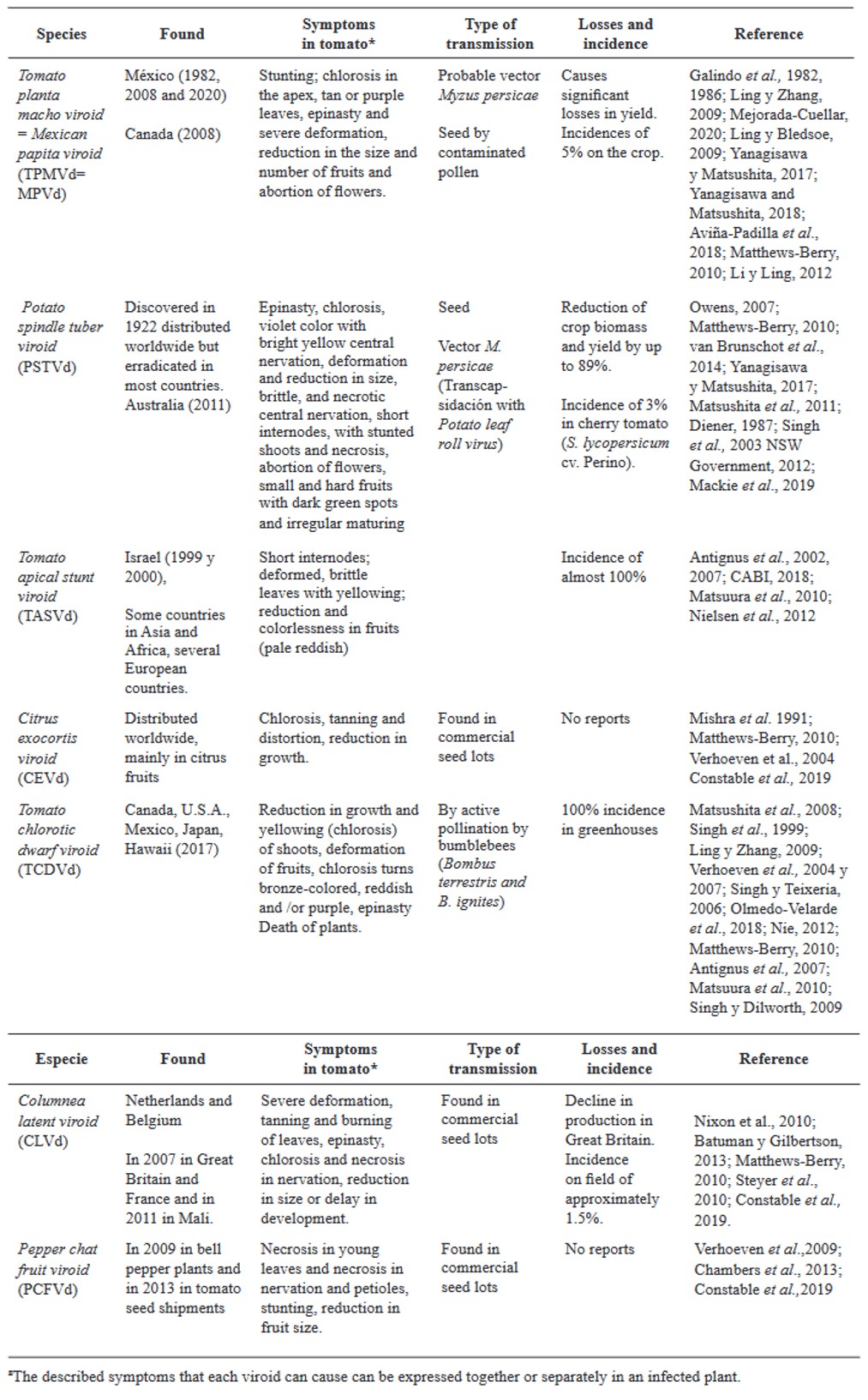
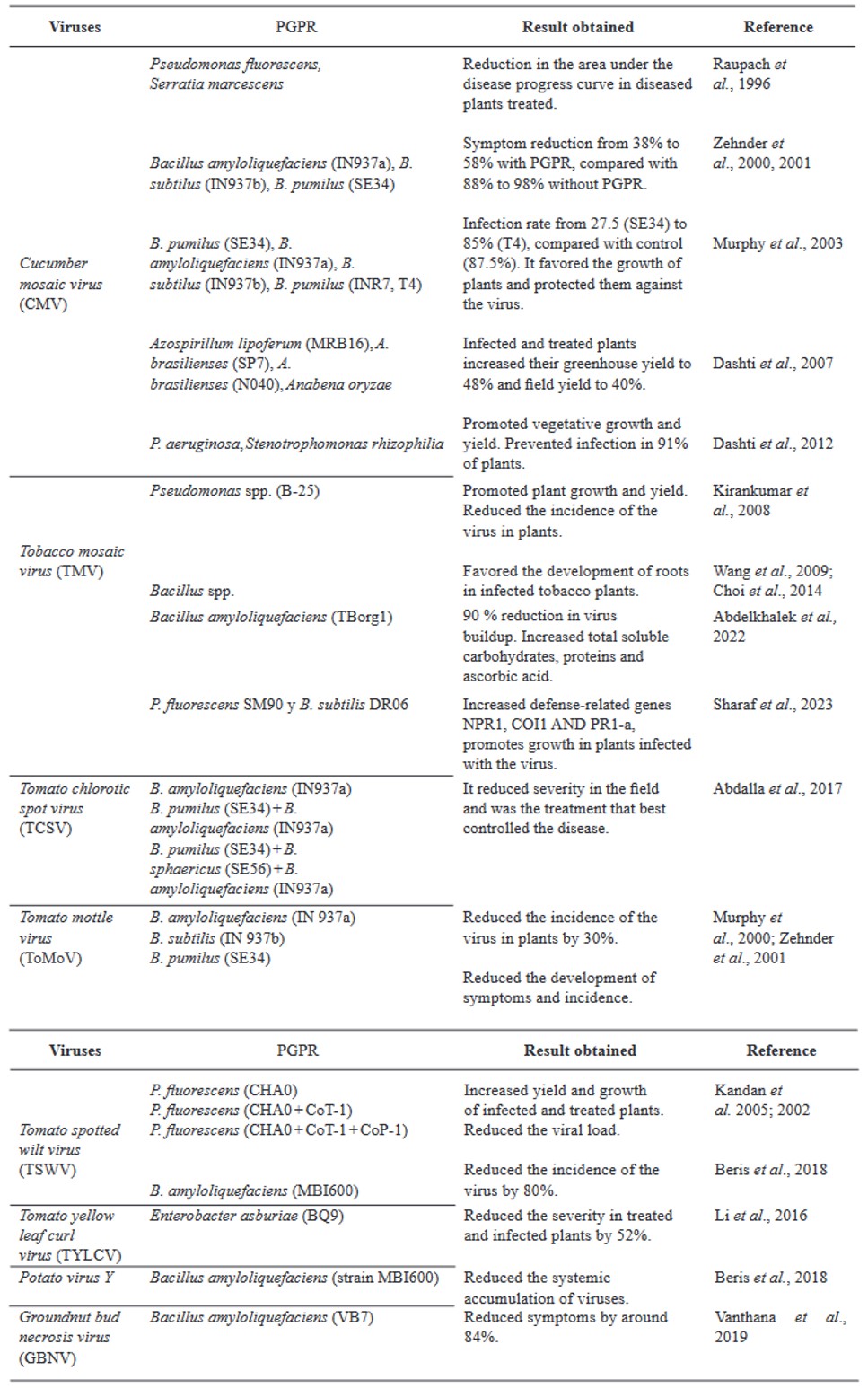
Resumen Los virus y viroides causan diversas enfermedades en jitomate (Solanum lycopersicum) en el mundo, generando pérdidas económicas importantes. Se han asociado alrededor de 312 virus y siete viroides, de los cuales más de 28 están presentes en México. Es necesaria la búsqueda de alternativas de manejo de estos fipatógenos, ya que la clásica eliminación de las primeras sintomáticas genera pérdidas de rendimiento, aunado a ello, la dificultad de evitar su diseminación. Por ello, el uso de rizobacterias promotoras del crecimiento vegetal (RPCV), puede ser una alternativa efectiva para el manejo de virus y viroides. Los géneros Pseudomonas, Bacillus, Azospirillum, Anabena y Stenotrophomonas, se han implementado contra virus reportados en jitomate: Cucumber mosaic virus (CMV), Tobacco mosaic virus (TMV), Tomato chlorotic spot virus (TCSV), Tomato mottle virus (ToMoV), Tomato spotted wilt virus (TSWV), Tomato yellow leaf curl virus (TYLCV), Potato virus Y (PVY), Groundnut bud necrosis virus (GBNV), con beneficios en la disminución de incidencia y severidad hasta un 80 % y un aumento de rendimiento superior al 40 %. En México solo se ha utilizado Bacillus. Se vislumbra que el uso de RPCV es una estrategía que podría mitigar el impacto de enfermedades virales y viroidales, que se puede integrar a un manejo integrado.
-
O copiar el link

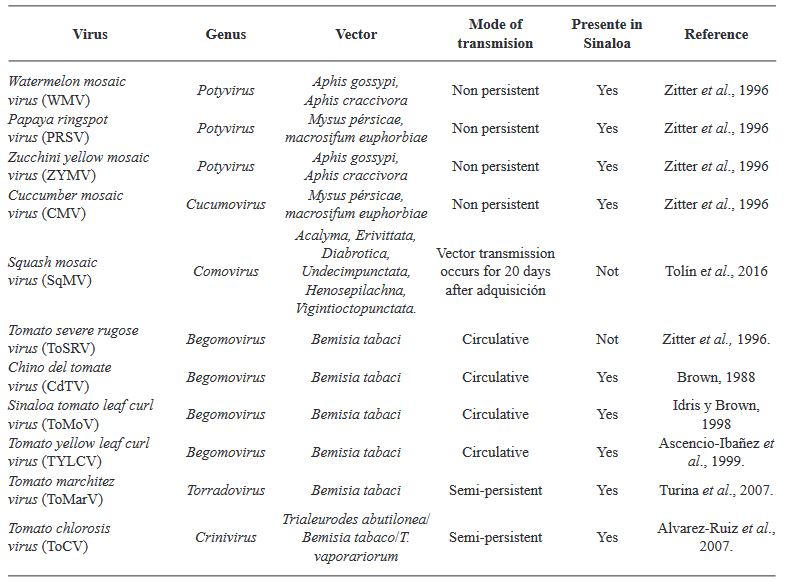
por Rubén Félix Gastélum, Gabriel Herrera Rodríguez, Karla Yeriana Leyva Madrigal, Guadalupe Arlene Mora Romero
Recibido: 31/7/2023 – Publicado: 28/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-4
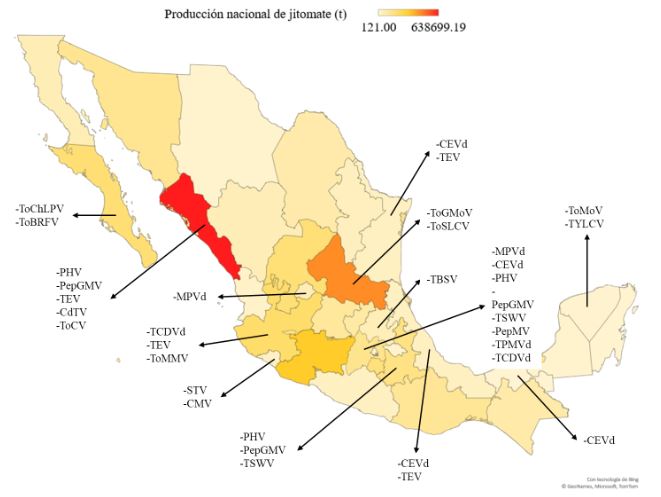
Resumen Se abordan especies de arvenses y ruderales de las familias Cucurbitaceae, Solanaceae en el norte de Sinaloa, como potenciales fuentes de inóculo para el desarrollo del Tomato apex necrosis virus (ToANV), Zucchini yellow mosaic virus (ZYMV), Watermelon mosaic virus (WMV), (Papaya ring spot virus (PRSV-W) y (Cucumber mosaic virus (CMV). Se hace referencia al girasol silvestre como potencial fuente de inóculo para la cenicilla (Golovinomyces spadiceus); se incluyen también el tabaquillo para el tizón foliar (Alternaria spp.), el chichiquelite para la mancha de la hoja (Curvularia moehlemvekiae), el zacate Johnson para el tizón foliar (Alternaria sp.), la higuerilla silvestre para el tizón foliar y el meloncillo silvestre para el mildiú (Pseudoperonospora cubensis). Se proponen líneas futuras de investigación multidisciplinarias enfocadas a la determinación de la patogenicidad en plantas cultivadas de virus y hongos asociados a plantas silvestres y viceversa. También se deberá estudiar la distribución espacio-temporal de plantas silvestres que pue den fungir como fuentes de inóculo, así como la de potenciales insectos vectores de enfermedades virales. La implementación de herramientas moleculares modernas, como la Secuenciación de Alto Rendimiento, para la detección de fitopatógenos es importante. Todo esto contribuirá a la aplicación de estrategias amigables con el ambiente para el control de las enfermedades en cultivos agrícolas en Sinaloa, en beneficio de los productores de hortalizas.
-
O copiar el link



El género Orthotospovirus en Costa Rica: Un caso centroamericano
por Mauricio Montero Astúa, Natasha Dejuk Protti, David Bermúdez Gómez, Elena Vásquez Céspedes, Laura Garita Salazar, Federico J. Albertazzi, Scott Adkins , Lisela Moreira Carmona
Recibido: 23/8/2023 – Publicado: 28/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-6
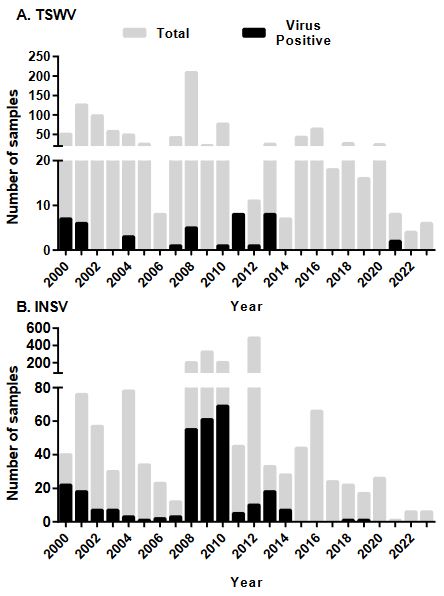
Resumen Antecedentes/Objetivo. El género Orthotospovirus, conocido por impacto significativo en una variedad de cultivos de importancia global, se manifiesta en virus emergentes con plantas económicamente perjudiciales. Aunque estos virus fitopatógenos están bien documentados en América del Norte y del Sur, su presencia y dinámica en América Central, particularmente en Costa Rica, un punto crítico entre los dos continentes, siguen siendo menos comprendidos. El objetivo de este trabajo fue determinar la prevalencia de los Orthotospovirus en Costa Rica y obtener secuencias parciales del genoma para analizar la variabilidad genética.
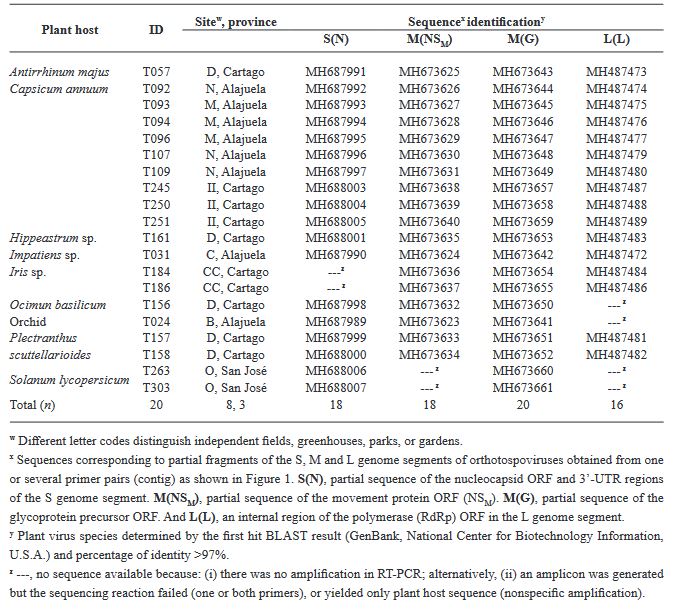
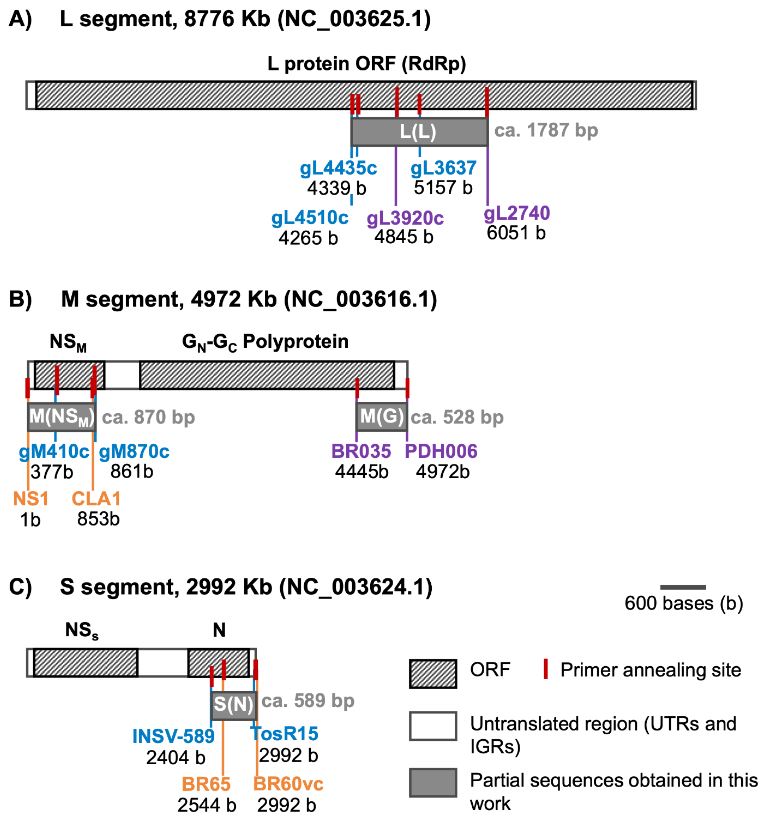
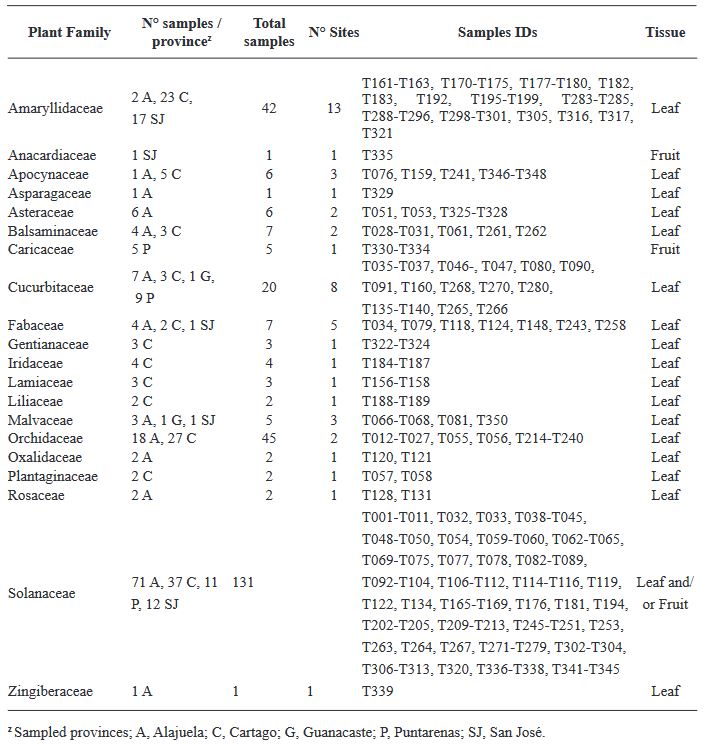
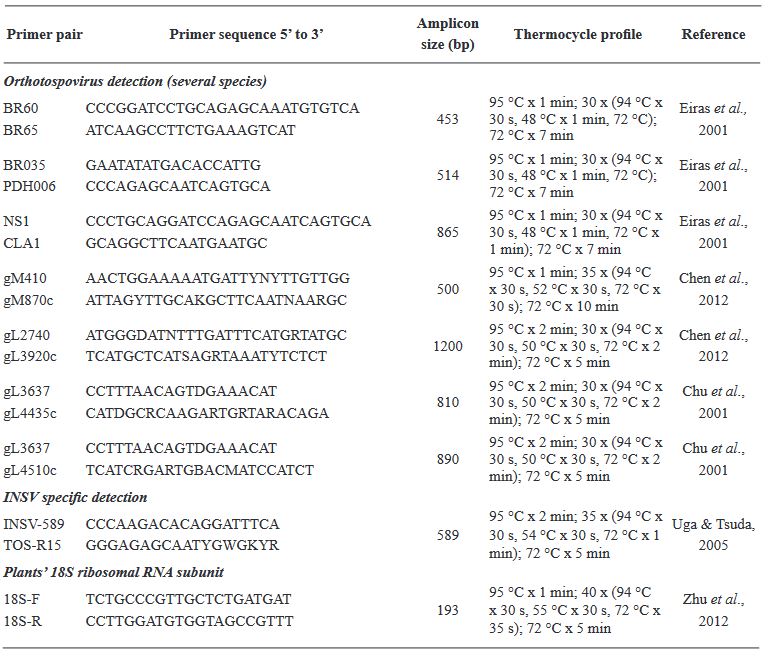
Materiales y Métodos. El estudio consistió en un análisis exhaustivo de 295 muestras de plantas utilizando el ensayo de inmunoadsorción ligado a enzimas (ELISA), una técnica sensible para detectar antígenos específicos, para evaluar la prevalencia de INSV, IYSV, TSWV y el serogrupo GRSV/TCSV. Una caracterización molecular más profunda de 20 muestras se realizó mediante la reacción en cadena de la polimerasa con transcripción inversa (RT-PCR), utilizando cebadores tanto de amplio espectro como específicos de especie para aumentar la precisión de la detección.
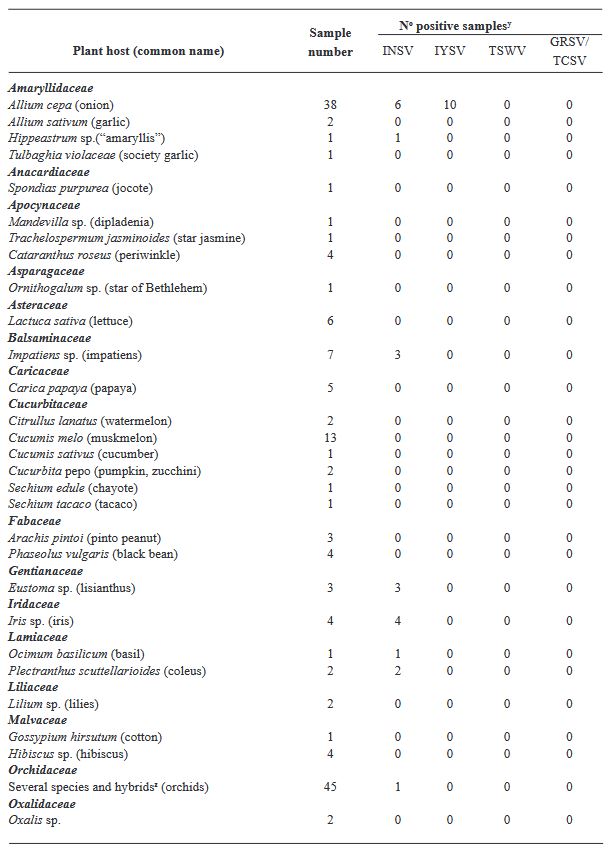
Resultados. Los resultados de ELISA revelaron la no detección de TSWV y el serogrupo GRSV/TCSV, divergiendo de informes de diagnóstico previos. La confirmación de INSV en Costa Rica a través de ELISA, RT-PCR y secuenciación parcial subraya su prevalencia tanto en campos abiertos como en invernaderos. A pesar de los informes diagnósticos anteriores que sugerían la presencia de TSWV en Costa Rica, nuestro estudio no detectó este virus. El análisis RT-PCR con cebadores degenerados tampoco encontró evidencia de otras especies de Orthotospovirus en nuestras muestras. La identificación de un haplotipo dominante de INSV, junto con tres variantes adicionales, sugiere la probabilidad de al menos dos introducciones independientes del virus en la región.
Conclusión. Estos hallazgos subrayan la necesidad de realizar muestreos e investigaciones más exhaustivas sobre los Orthotospovirus en América Central para comprender mejor su epidemiología e impacto en la agricultura
-
O copiar el link

![Figura 4. Gráfico de la Red de Haplotipos del Virus del anillado necrótico de la impatiens. Este gráfico representa una red de haplotipos inferida mediante el método TCS, basada en un alineamiento de secuencias de 261 bp del ORF de la proteína nucleocápside (segmento S[N]) del Virus del anillado necrótico de la impatiens. La red incluye datos de 81 aislados, representando visualmente las relaciones genéticas y la diversidad entre los diferentes haplotipos del virus.](img/RMF/Volumenes/NumEspeciales/VE4142023/RMF2023-6/Figure4.jpg)
![Figura 5. Análisis filogenético integral de aislados del virus del anillado necrótico de la impatiens. Esta figura muestra un árbol filogenético construido a partir de un alineamiento de secuencias concatenadas (2945 posiciones) que incluyen regiones del nucleocápside [S(N)], precursor de la glicoproteína [M(G)], proteína de movimiento [M(NSM)] y ORFs de la polimerasa viral [L(L)]. El árbol incluye aislados de varios países, identificados por códigos de tres letras según ISO 3166-1: CHN (China), ITA (Italia), KOR (Corea del Sur) y USA (Estados Unidos de América). Para los aislados de Costa Rica, códigos de letras adicionales y distintos entre paréntesis indican ubicaciones geográficas independientes. El análisis, realizado en MEGA X, utilizó el método de Máxima Verosimilitud con un modelo de parámetro Tamura-3 y una tasa de variación en nucleótidos distribuida gamma (+G), con 2000 permutaciones. La barra de escala indica el número de sustituciones de nucleótidos por sitio.](img/RMF/Volumenes/NumEspeciales/VE4142023/RMF2023-6/Figure5.jpg)
Viroma del nopal verdura en la zona centro de México
por Candelario Ortega Acosta, Daniel L. Ochoa Martínez, Reyna I. Rojas Martínez, Cristian Nava Díaz, Rodrigo A. Valverde
Recibido: 31/7/2023 – Publicado: 27/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-2
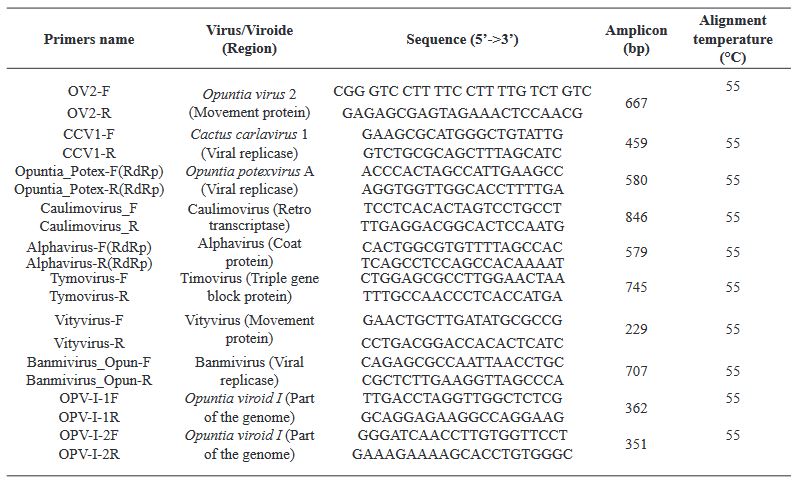
Resumen Antecedentes/Objetivo. En este estudio se aprovechó la capacidad de la secuenciación de alto rendimiento (HTS) para detectar virus en nopal verdura.
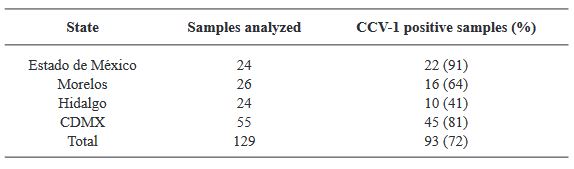
Materiales y Métodos. Se analizaron muestras del Estado de México (EDMX), Hidalgo y Morelos, así como de la Ciudad de México (CDMX).
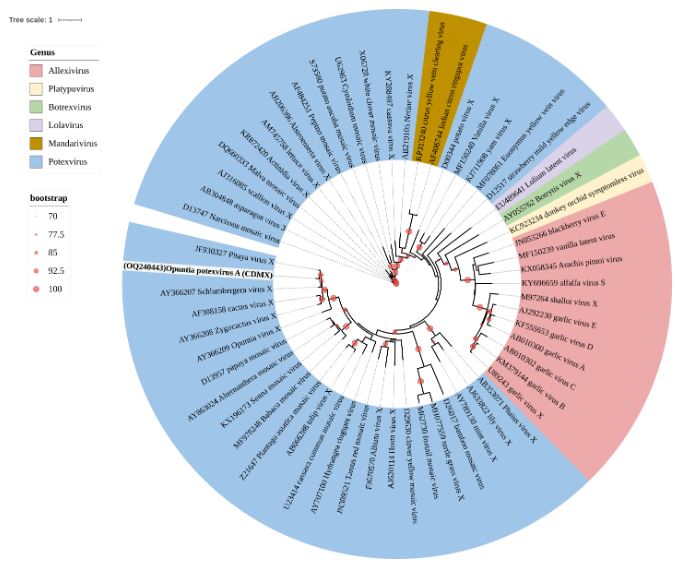
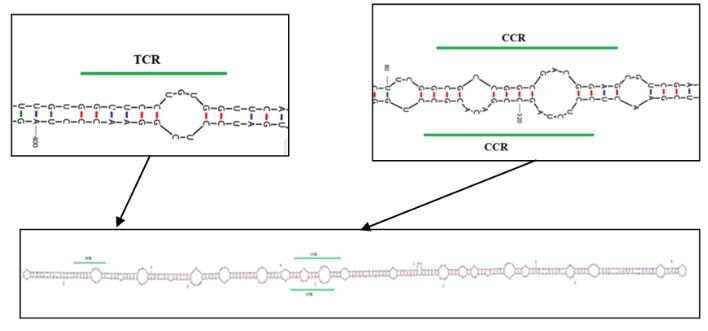
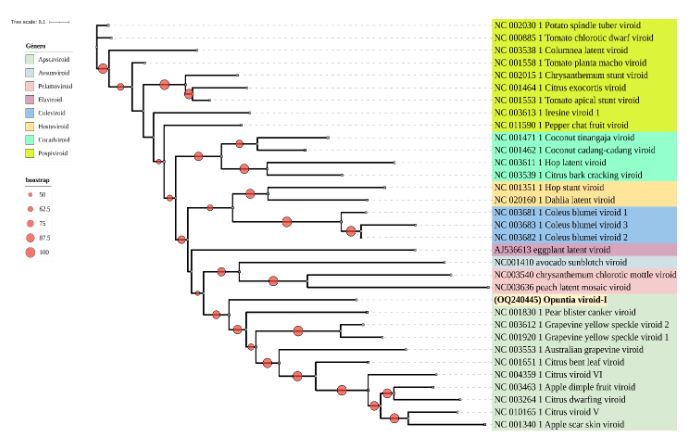
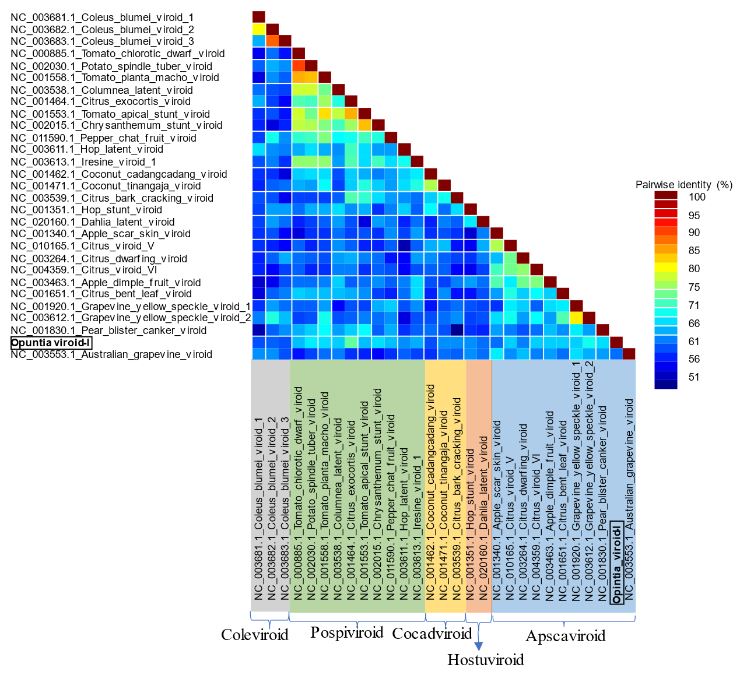
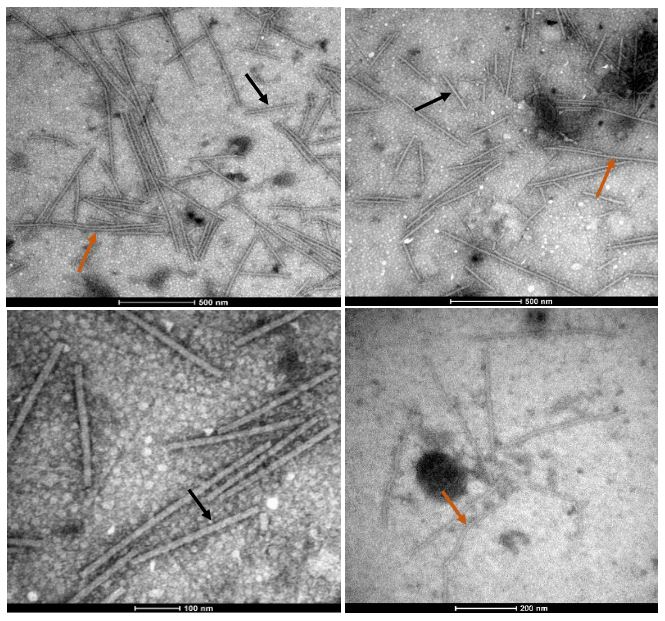
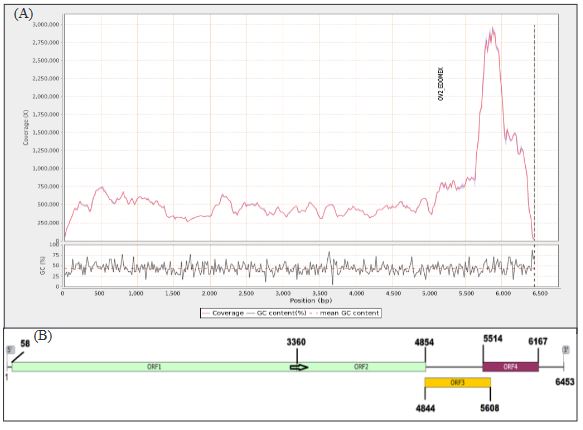
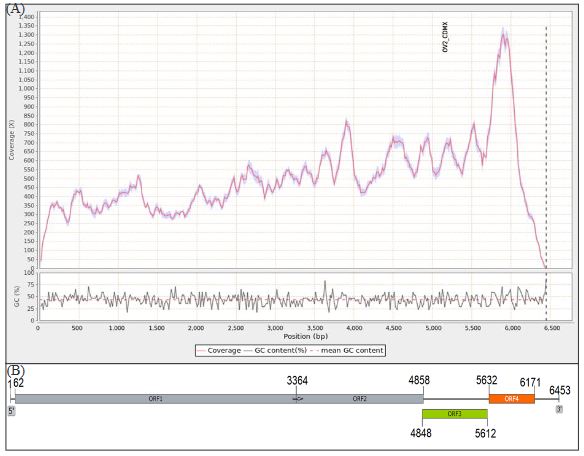
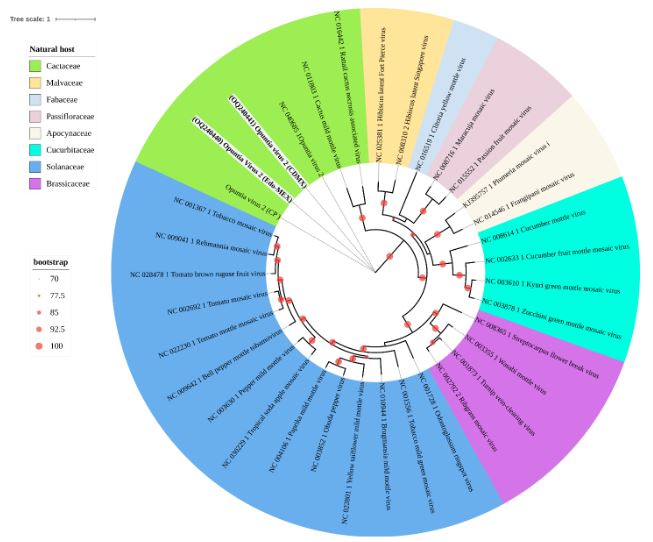
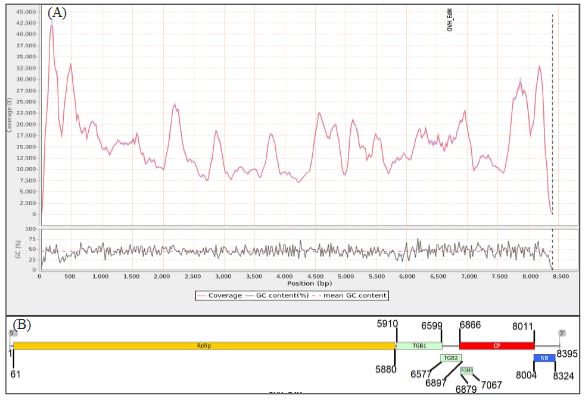
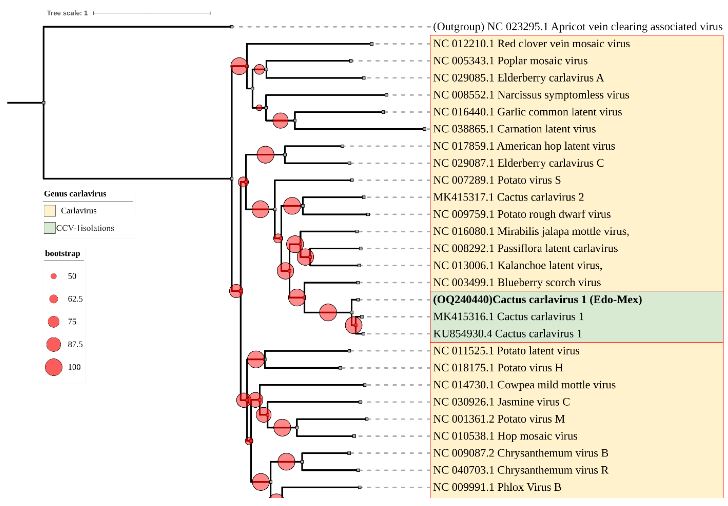
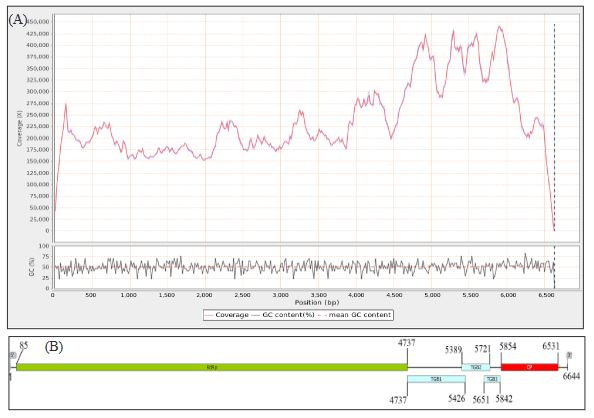
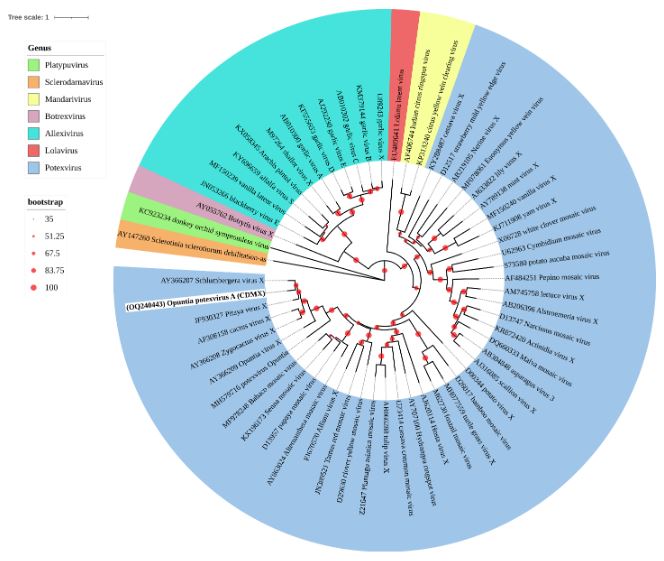
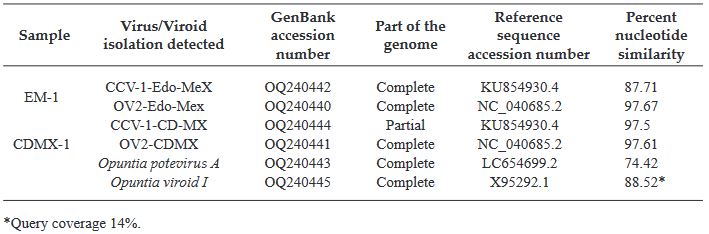
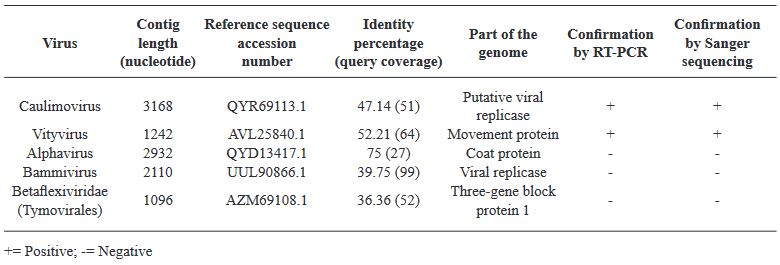
Resultados. En la muestra proveniente de EDMX, se detectaron y recuperaron los genomas de Opuntia virus 2 (OV2, género Tobamovirus) y Cactus carlavirus 1 (CCV-1, género Carlavirus). En la muestra proveniente de CDMX, además de OV2 y CCV-1, se detectó un nuevo viroide y un potexvirus. El primero tiene un genoma RNA circular con 412 nt de longitud para el cual se propone el nombre de “Opuntia viroid I” (OVd-I). La estructura primaria de este viroide mostró una identidad de secuencia de nucleótidos de menos del 80 % con cualquiera de los viroides actualmente conocidos y una relación filogenética con el género Apscaviroid, (Familia Pospiviroidae) con el que comparte motivos estructurales conservados.
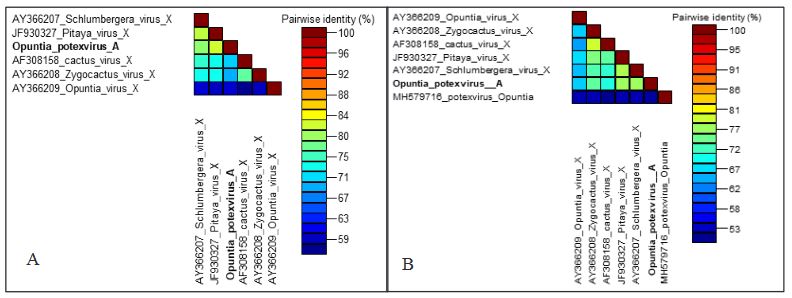
Conclusión. El nuevo potexvirus se denominó Opuntia potexvirus A (OPV-A), cuya secuencia de la replicasa viral tiene un 77.7 % de identidad de aminoácidos con Schlumbergera virus X. Finalmente, en 93 (72 %) de 129 muestras de nopal verdura recolectadas en las cuatro entidades, se detectó al CCV-1.
-
O copiar el link