-
O copiar el link


Filtros de Contribuciones
Buscar Contribución:
por Dulce Jazmín Hernández Melchor, Ronald Ferrera Cerrato, Clemente de Jesús García Ávila, Alejandro Alarcón
Recibido: 09/7/2023 – Publicado: 29/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2307-2
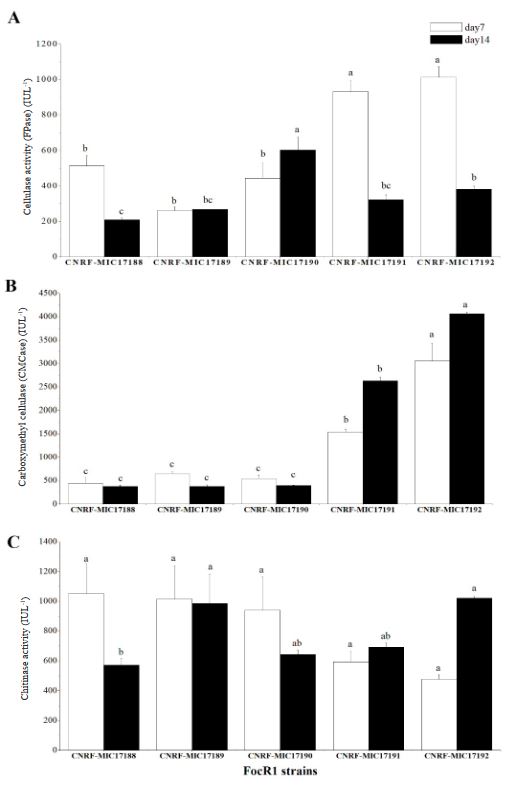
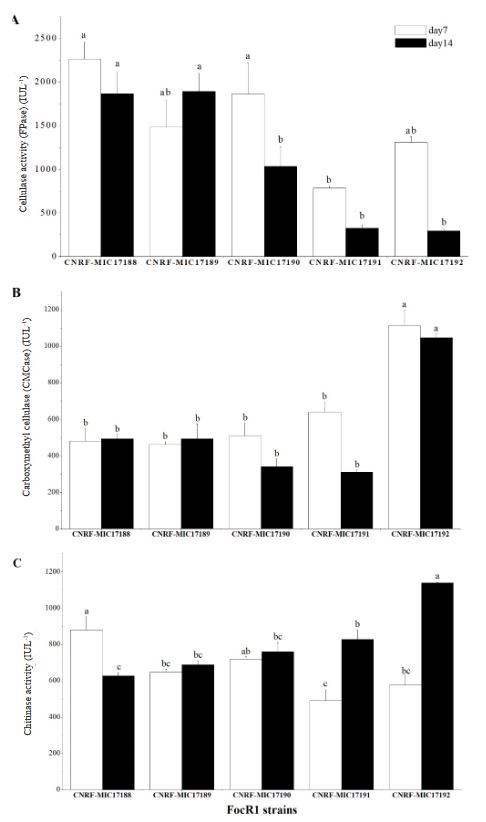
Resumen Antecedentes/Objetivo. Fusarium tiene la capacidad de producir enzimas hidrolíticas de interés en la industria de los alimentos o alcohólica para descomponer compuestos orgánicos naturales. Este trabajo estudió la capacidad de Fusarium oxysporum f.sp. cubense raza 1 (FocR1) para producir enzimas celulasas y Quitinasas en cultivo sumergido utilizando diferentes fuentes de carbono.
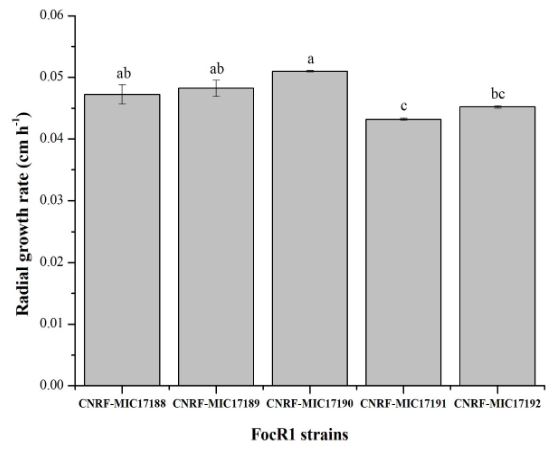
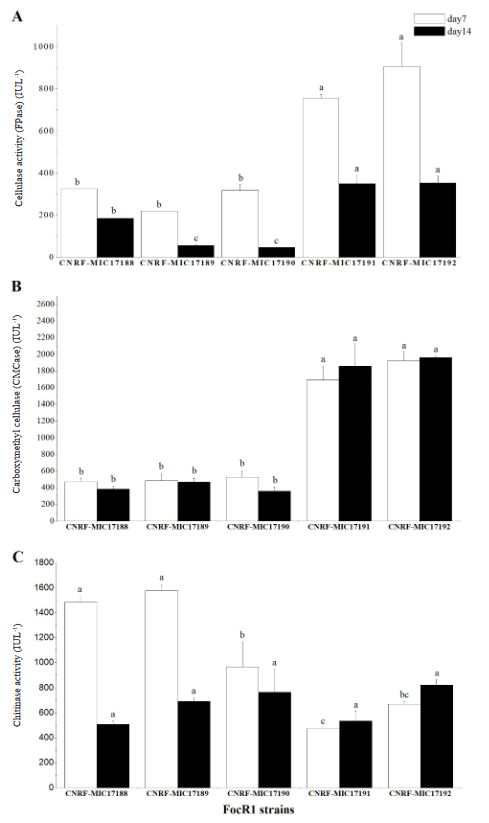
Materiales y Métodos. Cinco cepas de FocR1 (CNRF-MIC17188, CNRFMIC17189, CNRF-MIC17190, CNRF-MIC17191, y CNRF-MIC17192) se utilizaron en cultivo sumergido para la degradación de tres sustratos [papel filtro, papel periódico, y Quitina (Sigma®)], evaluando la velocidad de crecimiento radial (VCr) y la actividad enzimática cuantitativa (FPase, CMCase y Quitinasa).
Resultados. La VCr en las cinco cepas de FocR1 osciló en un rango de 0.043 a 0.051 cm h-1. A los días 7 y 14, las cinco cepas de FocR1 produjeron celulasas y Quitinasas al utilizar los tres sustratos. De acuerdo con el análisis estadístico, las cepas CNRF-MIC17191 y CNRF-MIC17192 presentaron los mejores resultados de actividades enzimáticas. Conclusiones. Las cinco cepas de FocR1 pueden utilizarse como una fuente comercial de celulasas y Quitinasas, así como ser candidatas potenciales para bioconvertir complejas fuentes de carbono para su futuro aprovechamiento en procesos industriales.
-
O copiar el link
Escala diagramática para cuantificar la severidad de mancha café en el cultivo de haba
por Ernesto Alonso López Reyes, Álvaro Castañeda Vildózola, Jesús Ricardo Sánchez Pale, Alejandra Contreras Rendón, Juyma Mayvé Fragoso Benhumea, Rómulo García Velasco
Recibido: 15/9/2022 – Publicado: 26/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2209-4
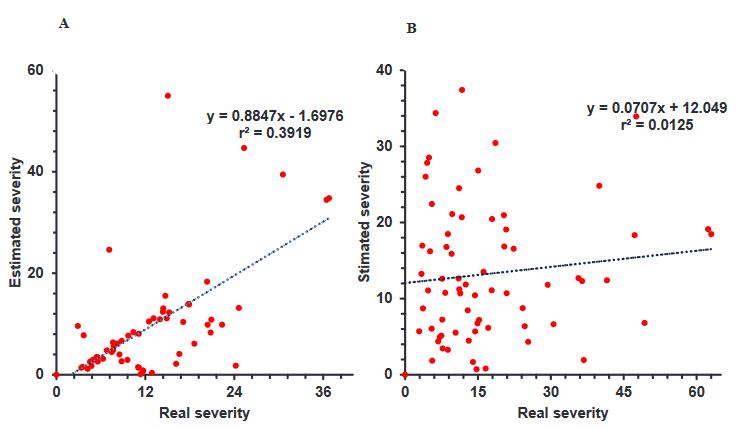
Resumen Antecedentes/Objetivo. El objetivo de esta investigación fue diseñar y validar una escala diagramática de severidad de la mancha café en haba.
Materiales y Métodos: En tres plantaciones comerciales, se recolectaron 120 foliolos con diferente nivel de daño de mancha café, realizando una selección visual a partir de la sintomatología. Se digitalizaron 60 foliolos para evaluarse con el software APS PRESS ©Assess 2.0 y determinar el valor de la severidad real de cada foliolo.
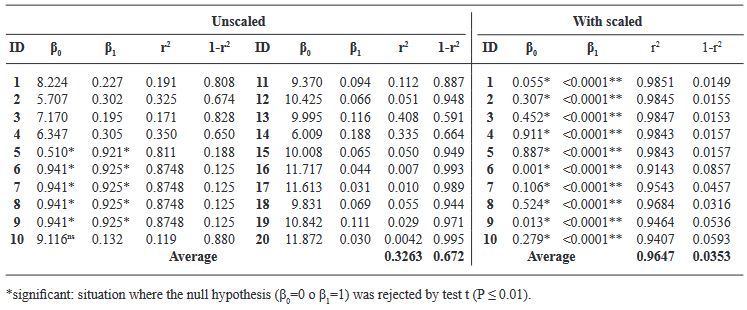
Resultados. Los valores de severidad permitieron generar una escala diagramática conformada por seis clases diferentes 0(0.0), 1(0.1-6.0), 2(6.1-10.0), 3(10.1-15.0), 4(15.1-40.0), 5(> 40.1-100). 20 evaluadores sin experiencia realizaron la primera evaluación visual (sin escala diagramática) de los foliolos con diferentes grados de daño. Posteriormente, se realizó una segunda evaluación, con 10 evaluadores, con apoyo de la escala. Los resultados obtenidos de cada evaluador se analizaron mediante una regresión lineal simple obteniendo valores de r2= 0.0042 a 0.8748, β0 de 0.51 a 9.11, y β1 de 0.132 a 0.925, sin uso de escala. Con uso de escala se obtuvieron valores de r2= 0.9143 a 0.9851, β0 de 0.001 a 0.911 y β1<0.001.
Conclusión. La escala diagramática de severidad generada fue validada y reproducible, mostrando una alta confiabilidad
-
O copiar el link

por Juanita Guadalupe Hollman Aragón, Mirella Romero Bastidas, Pablo Misael Arce Amezquita, Alejandro Palacios Espinosa
Recibido: 15/3/2023 – Publicado: 19/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2303-1
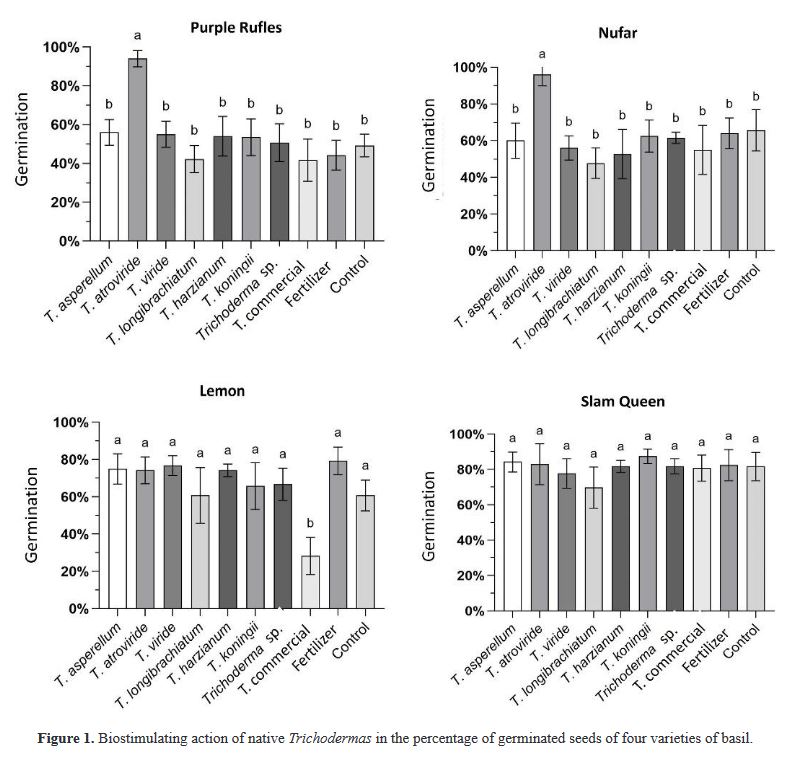
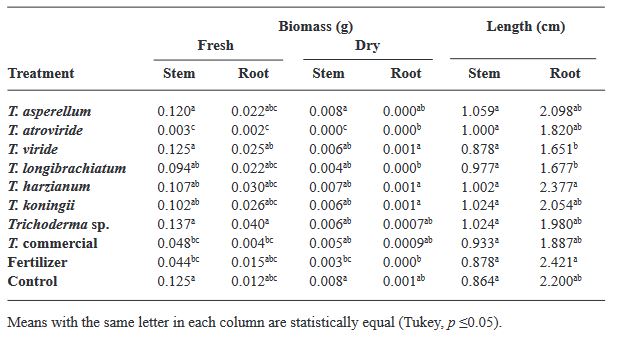
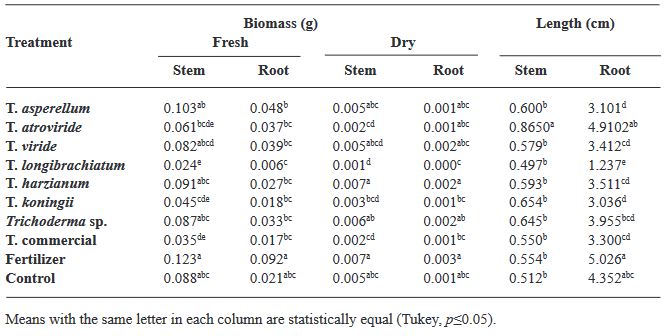
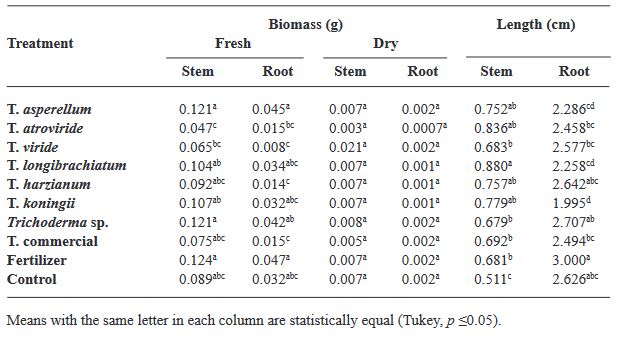
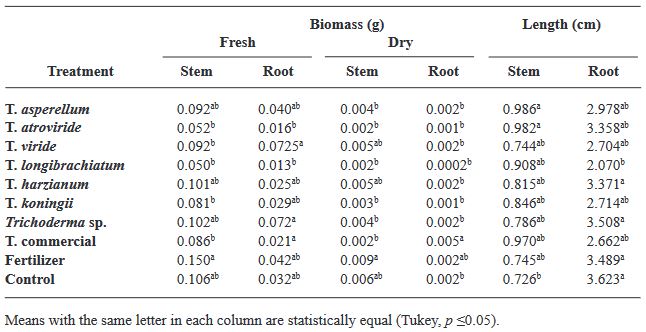
Resumen Antecedentes/Objetivo. Trichoderma es una herramienta eficiente como bioestimulante en cultivo de albahaca. Sin embargo, solo algunas especies han sido estudiadas sobre cultivares específicos. Por lo anterior, el objetivo de esta investigación fue evaluar la eficacia bioestimulante de cepas nativas de Trichoderma sobre la germinación y crecimiento de cuatro variedades de albahaca.
Materiales y Métodos. En el estudio se utilizaron seis especies de Trichoderma (T. asperellum, atroviride, viride, longibrachiatum, harzianum, koningii y Trichoderma sp.), una cepa de Trichoderma comercial (T. harzianum), fertilizante sintético (T17) y el control. 30 semillas de las variedades Purple Ruffles, Lemon, Siam Queen y Nufar fueron tratadas con una suspensión de esporas de cada Trichoderma. 48 h después, las semillas se sembraron e incubaron a 28 °C con un fotoperiodo de 12 h luz/oscuridad. Las variables a evaluar fueron; Tasa y porcentaje de germinación, biomasa y longitud de plántulas.
Resultados. T. atroviride presentó el mayor efecto bioestimulante en germinación. Mientras que T. asperellum registró una eficacia incrementada en biomasa y longitud de la planta en las cuatro variedades. La acción del T. comercial fue menor en todos los casos.
Conclusión. Este estudio demuestra que las cepas nativas de Trichoderma poseen efecto bioestimulante en las plantas y presentan mayor eficacia que las especies de tipo comercial.
-
O copiar el link

por Daniel Castrillo Sequeira, Rodrigo Jiménez Robles, Milagro Granados Montero
Recibido: 18/9/2023 – Publicado: 27/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2309-3
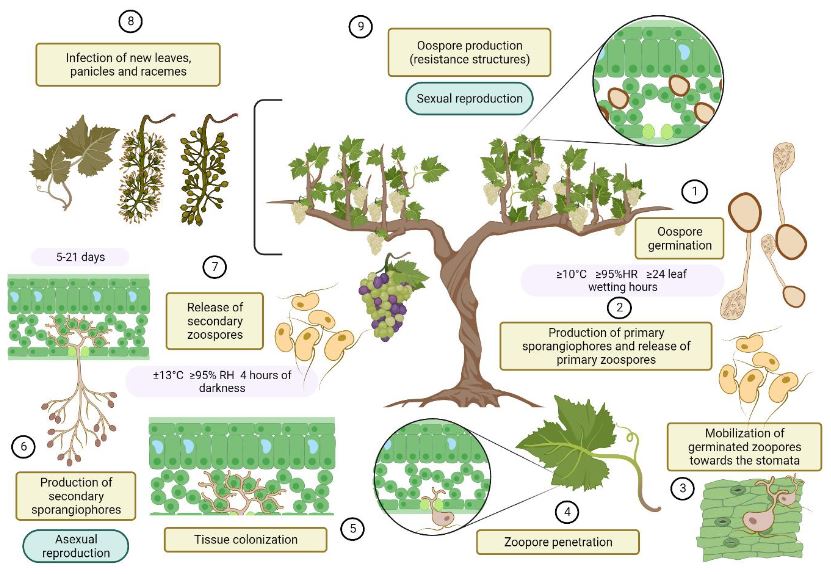
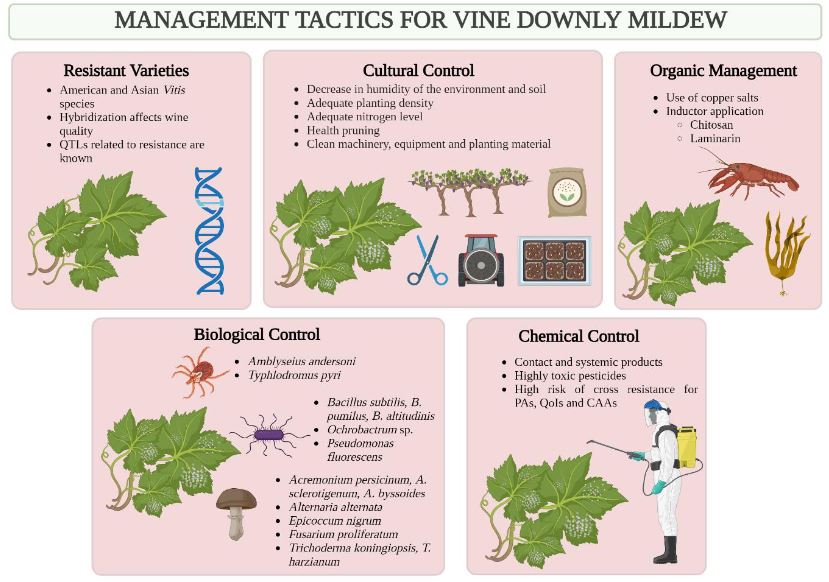
Resumen La viticultura es una de las actividades agrícolas de mayor antigüedad, y su explotación tradicionalmente se ha limitado a zonas de clima templado, de donde la vid (Vitis vinifera) y el vino son originarios. Ante los efectos del cambio climático, cada vez más áreas pierden capacidad para el desarrollo de este cultivo, y los trópicos se presentan como regiones potenciales para este mercado. En Costa Rica, la actividad vitícola se ha reportado desde mediados del siglo XX, sin embargo, la información técnica sobre el cultivo es escasa. El mildiú velloso, causado por el oomicete Plasmopara viticola, representa una de las enfermedades de mayor impacto económico para la viticultura a nivel global, así como el problema fitosanitario más limitante en Costa Rica. En condiciones de alta humedad, el desarrollo del patógeno es acelerado, y el hospedante se mantiene susceptible durante todo el ciclo de cultivo, lo cual dificulta el manejo adecuado de las epidemias. Globalmente, el combate químico es la estrategia de manejo más común; sin embargo, la aparición de poblaciones de P. viticola con resistencia a fungicidas se ha observado en la mayoría de las zonas productoras, por lo que la búsqueda de alternativas más ecológicas constituye una necesidad. En la actualidad, Costa Rica no cuenta con estrategias de manejo integrado que permitan la producción sostenible, y solo existe un producto registrado para la protección contra este patógeno. Esta situación justifica prestar mayor atención al estudio de este patosistema.
-
O copiar el link

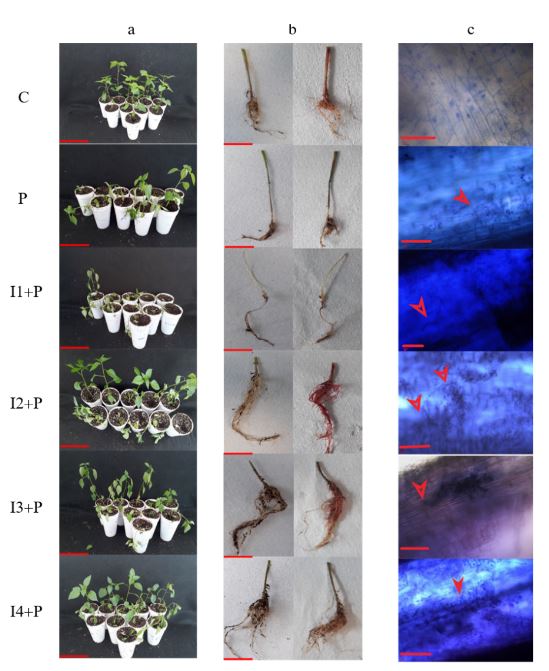
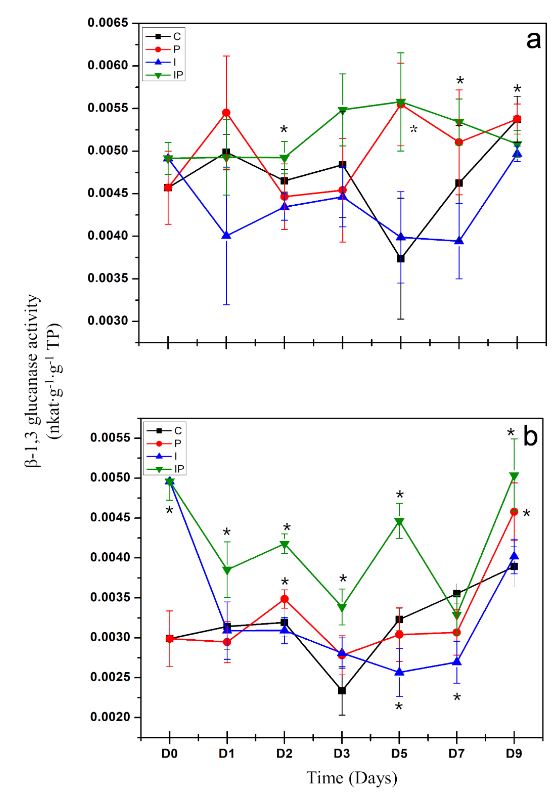
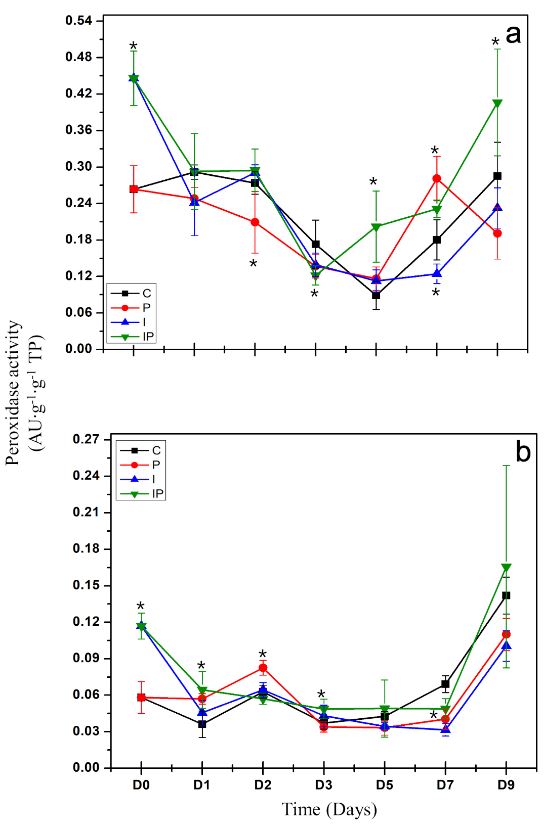
Inducción de respuesta de defensa por inulina de tubérculos de dalia (Dahlia sp.) en Capsicum annuum
por Julio César López Velázquez, Soledad García Morales, Joaquín Alejandro Qui Zapata, Zaira Yunuen García Carvajal, Diego Eloyr Navarro López, Rebeca García Varela
Recibido: 10/5/2023 – Publicado: 29/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2305-2
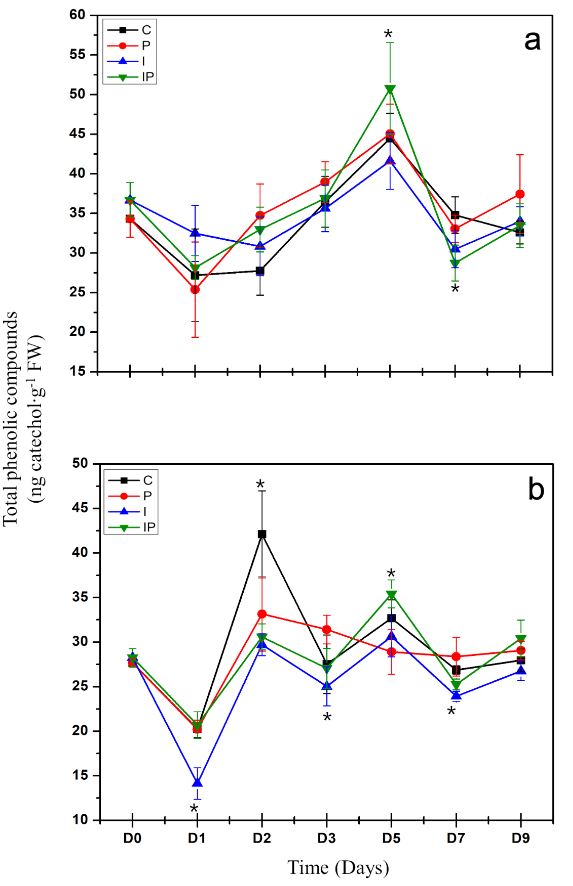
Resumen Antecedentes/Objetivo. Phytophthora capsici es el agente causal de la marchitez del chile. Entre las estrategias para su control está el uso de inductores de resis tencia. Los fructanos son moléculas que presentan propiedades biológicas interesantes, incluyendo la capacidad de inducir mecanismos de resistencia en algunas plantas. En este trabajo se evaluó el efecto de protección de cuatro concentraciones de inulina de tubérculos de dalia en chile infectado con P. capsici.
Materiales y Métodos. La concentración que presentó mayor protección se eligió para evaluar la inducción de respuesta de defensa mediante la actividad enzimática de β-1,3 glucanasas, peroxidasas y la producción de compuestos fenólicos totales.
Resultados. La inulina mostró un efecto protector contra la infección a concentraciones de 100 a 300 μM, ya que disminuyeron los síntomas y las plántulas mostraron mejor desarrollo vegetativo. Se observó que la inulina en concentración 200 μM indujo una respuesta de defensa efectiva asociada al aumento de la actividad de β-1,3 glucanasas y peroxidasas mediante una respuesta local y sistémica en las plántulas. Esta respuesta fue diferenciada entre las plántulas tratadas con inulina y las plántulas infectadas con P. capsici.
Conclusión. Se concluyó que la inulina tiene la capacidad de proteger al chile de P. capsici por la inducción de resistencia.
-
O copiar el link
por María Guadalupe Aguilar Rito, Amaury Martín Arzate Fernández, Hilda Guadalupe García Núñez, Tomas Héctor Norman Mondragón
Recibido: 02/10/2023 – Publicado: 20/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-1
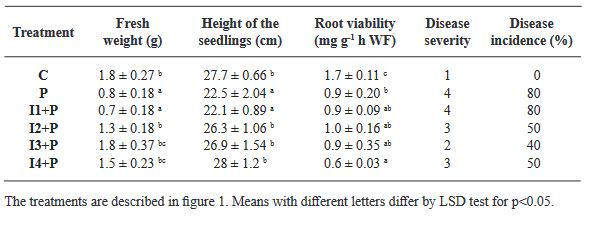
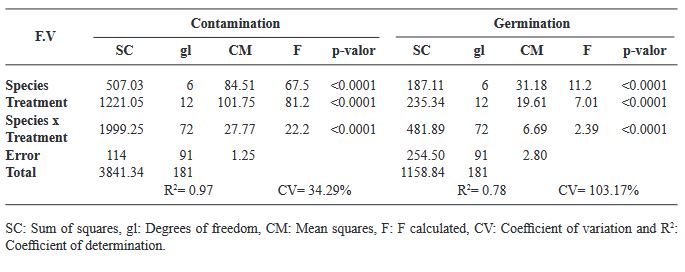
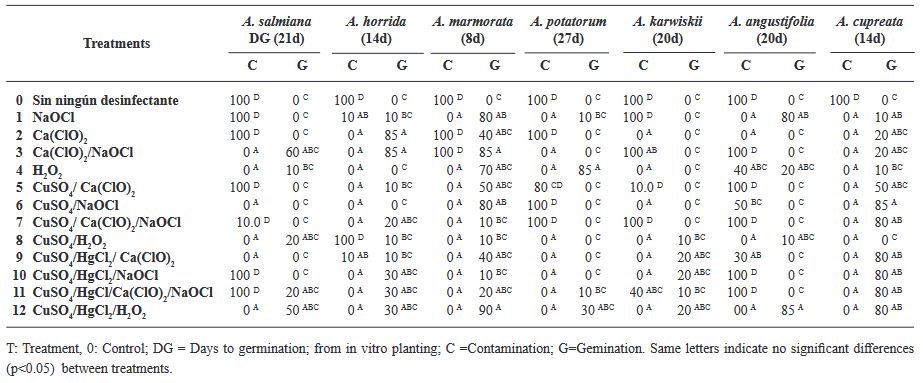
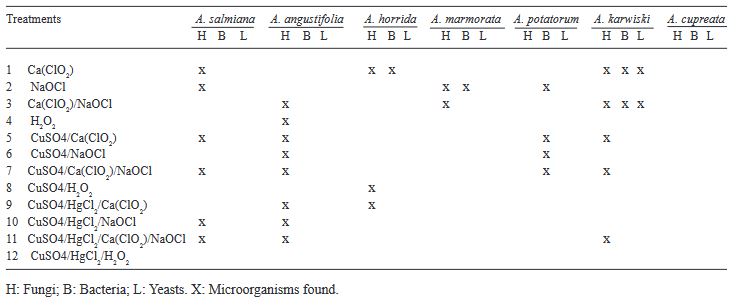
Resumen Antecedentes/Objetivo. La desinfección de las semillas de Agave es un paso crucial en el cultivo in vitro para prevenir la contaminación, la cual puede ser causada por microorganismos como bacterias, hongos y virus que pueden afectar el crecimiento de las plántulas y reducir la tasa de germinación de las semillas. Por lo tanto, la desinfección adecuada de las semillas es esencial para garantizar un crecimiento vegetal vigoroso y saludable. El objetivo principal de esta investigación fue: evaluar 12 tratamientos para generar un protocolo eficiente de desinfección in vitro en semillas de seis especies de Agave mezcalero y un pulquero con desinfectantes y combinaciones diferentes.
Materiales y Métodos. Los desinfectantes utilizados fueron; Peróxido de Hidrógeno al 3 % por 24 h, Hipoclorito de Sodio comercial al 5 % (v/v) por 5 min, Hipoclorito de Calcio al 8 % (p/v) por 15 min, Sulfato de cobre (PENTAMAX®) al 30 % (v/v) por 10 min, Cloruro de Mercurio II al 0.1 % (p/v) por 10 min.
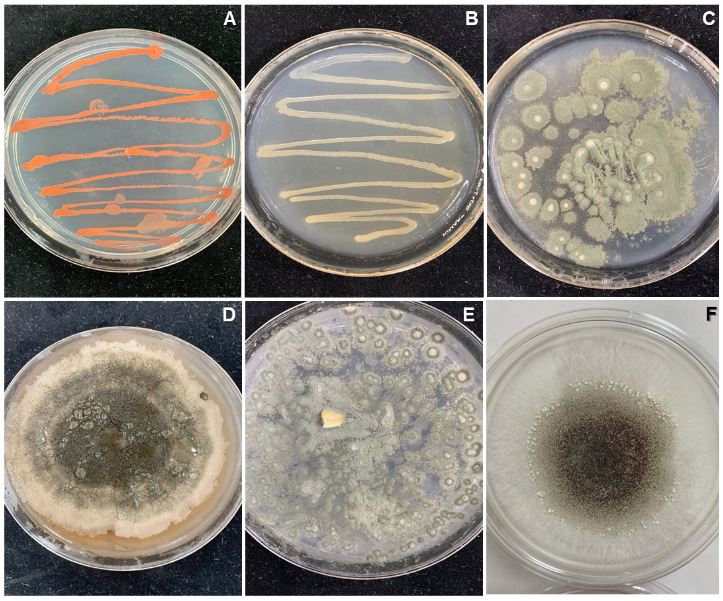
Resultados. Los microorganismos contaminantes identificados fueron cuatro géneros de hongos: Monilinia sp., Aspergillus sp., Penicillium sp., y Alternaria alternata, una bacteria; Bacillus sp., y una levadura, Schizosaccharomyces sp.
Conclusión. El mejor tratamiento para la desinfección de las semillas fue Peróxido de Hidrógeno al 3 % por 24 h en combinación con Sulfato de cobre al 30 % (v/v) por 10 min y Cloruro de Mercurio II al 0.1 % (p/v) por 10 min, obteniendo un 100 % de desinfección y logrando que germinaran todas las especies.
-
O copiar el link

por Edgar Guevara Avendaño, Itzel Anayansi Solís García, Alfonso Méndez Bravo, Fernando Pineda García, Guillermo Angeles Alvarez, Carolina Madero Vega, Sylvia P. Fernández Pavía, Alejandra Mondragón Flores, Frédérique Reverchon
Recibido: 17/9/2023 – Publicado: 08/12/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2309-2
Resumen Antecedentes/Objetivo. El objetivo fue corroborar la capacidad como agente de biocontrol de la cepa Bacillus sp. A8a en árboles de aguacate (Persea americana) infectados por Phytophthora cinnamomi.
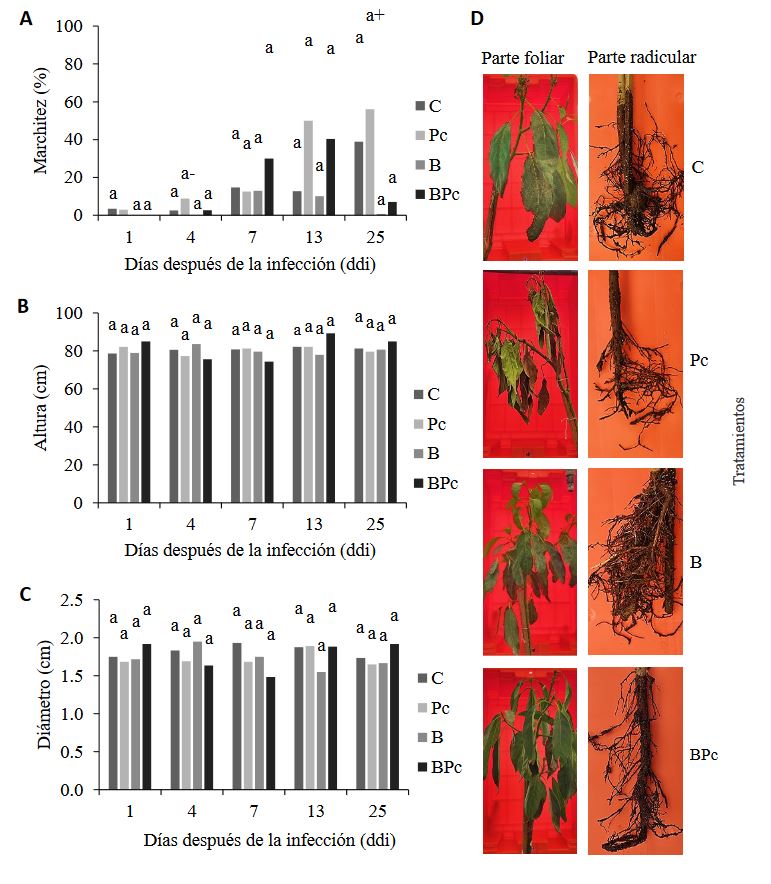
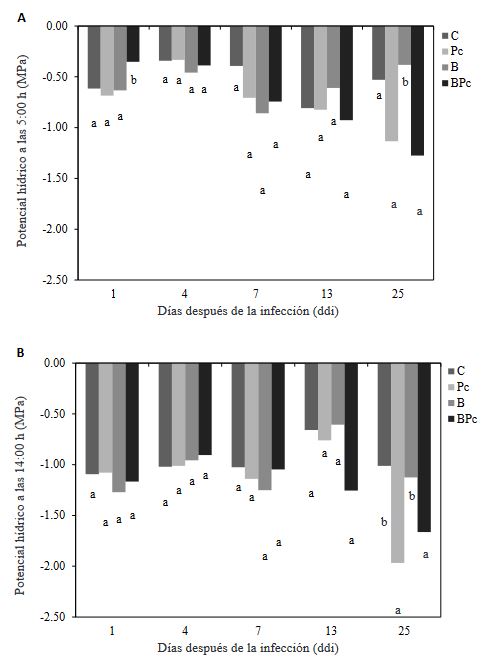
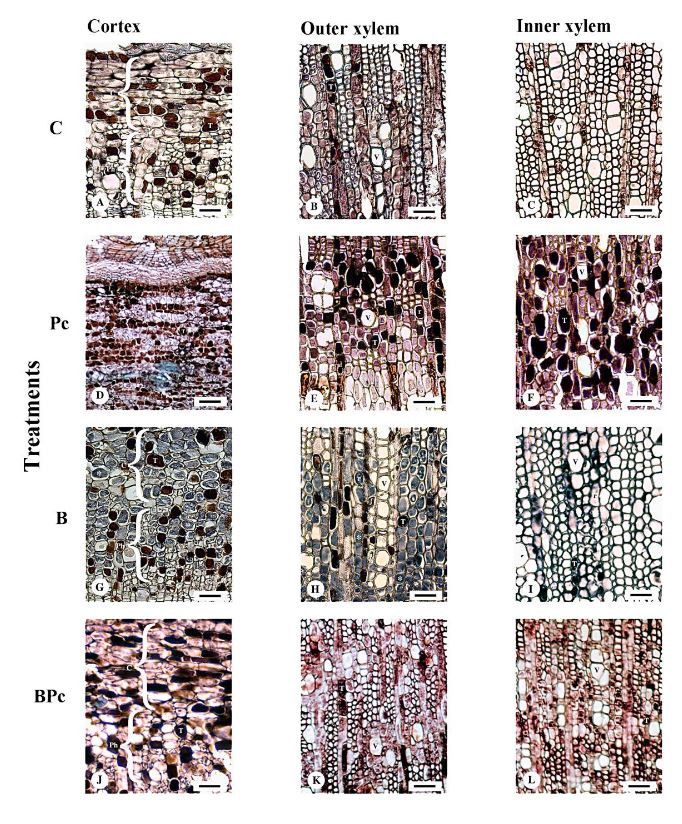
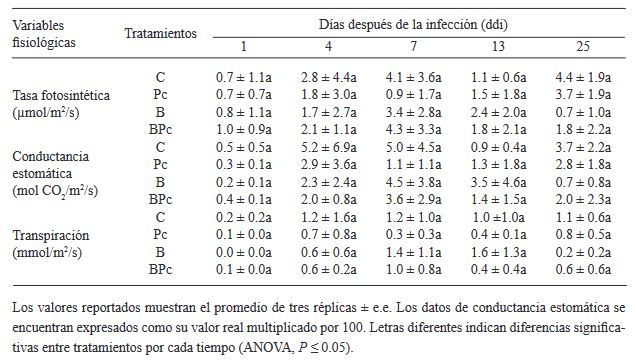
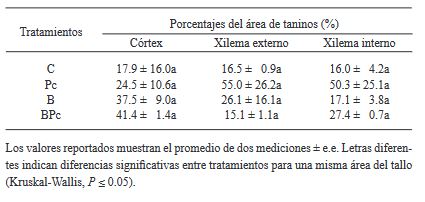
Materiales y Métodos. Se implementó un experimento en invernadero con cuatro tratamientos: 1) plantas control; 2) plantas infectadas por P. cinnamomi; 3) plantas inoculadas con Bacillus sp. A8a; 4) plantas infectadas por P. cinnamomi e inoculadas con Bacillus sp. A8a. Se evaluaron distintas variables morfo-fisiológicas durante el experimento, hasta 25 días después de la infección (ddi). Además, se analizó la densidad de taninos en cortes de tallo a los 25 ddi, para determinar la respuesta de defensa de los árboles ante la enfermedad.
Resultados. La inoculación de la cepa A8a redujo los síntomas de marchitez en un 49% a los 25 ddi, en comparación con las plantas no inoculadas. No se encontraron diferencias en las variables morfo-fisiológicas registradas entre tratamientos. Sin embargo, se detectó una mayor acumulación de taninos en el xilema de tallos de plantas infectadas, mientras que las plantas inoculadas con la cepa A8a mostraron una mayor densidad de taninos en el córtex.
Conclusión. Nuestros resultados confirman la actividad de biocontrol de Bacillus sp. A8a en aguacate e indican que la acumulación diferencial de taninos en el córtex de plantas inoculadas con la bacteria podría contribuir a la mayor tolerancia de las plantas de aguacate ante la marchitez por Phytophthora
-
O copiar el link
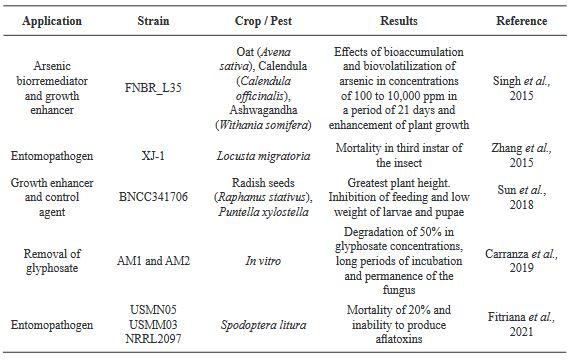
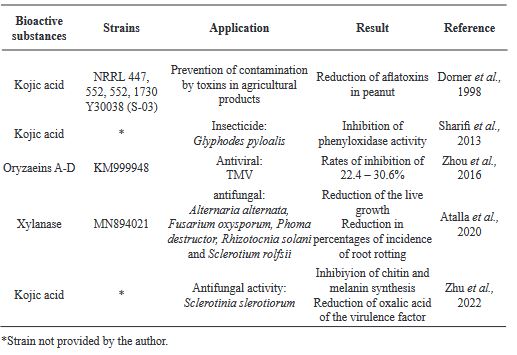
Aspergillus oryzae: Una oportunidad para la agricultura
por Karen Berenice García Conde, Ernesto Cerna Chávez, Yisa María Ochoa Fuentes, Jazmín Janet Velázquez Guerrero
Recibido: 23/2/2023 – Publicado: 14/11/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2302-2
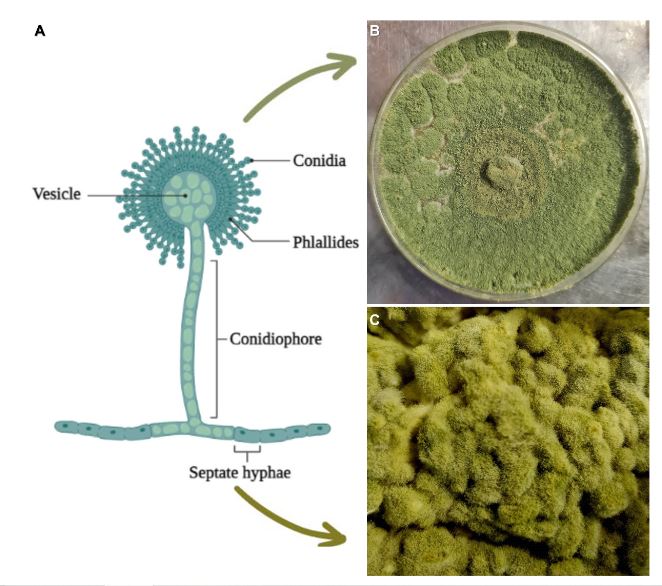
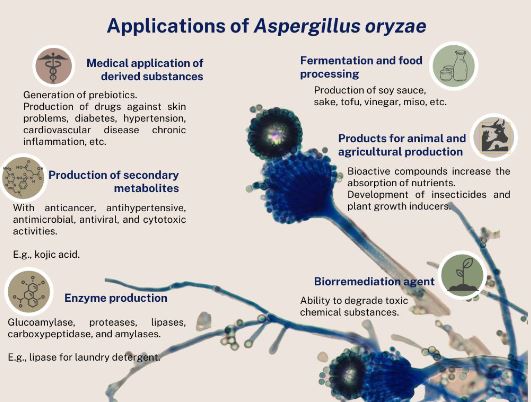
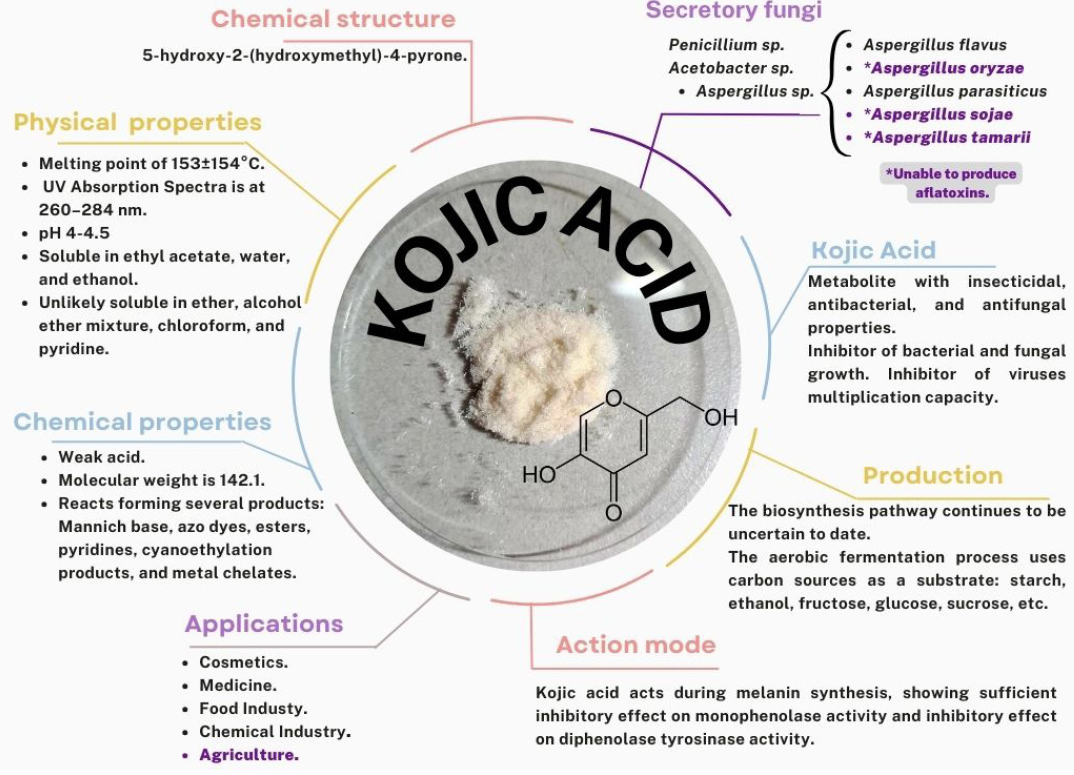
Resumen Aspergillus oryzae es un hongo filamentoso, capaz de degradar diversas sustancias empleando enzimas, por lo que es ampliamente utilizado en la industria biotecnológica, productos farmacéuticos, enzimas para uso industrial, agentes blanqueadores y tratamientos textiles anticontaminación. Sin embargo, son pocos los trabajos centrados en las aplicaciones de campo de este microorganismo. El presente manuscrito revisa las posibles aplicaciones con potencial benéfico de A. oryzae y algunos subproductos en la agricultura como control biológico, inductor de crecimiento y biorremediador de suelos contaminados con metales pesados.
-
O copiar el link
por Juan Agustin Gonzalez Cruces, José Sergio Sandoval Islas, Cristian Nava Díaz, Maricarmen Sandoval Sánchez
Recibido: 30/11/2023 – Publicado: 16/11/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2211-1
Resumen Antecedentes/Objetivo. El objetivo de esta investigación fue identificar el agente causal de la pudrición del rizoma del espárrago, así como evaluar distintos métodos de inoculación y la severidad de los aislados.
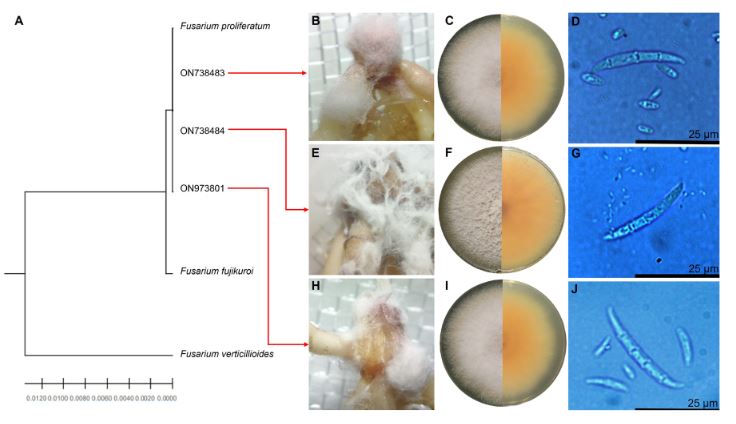
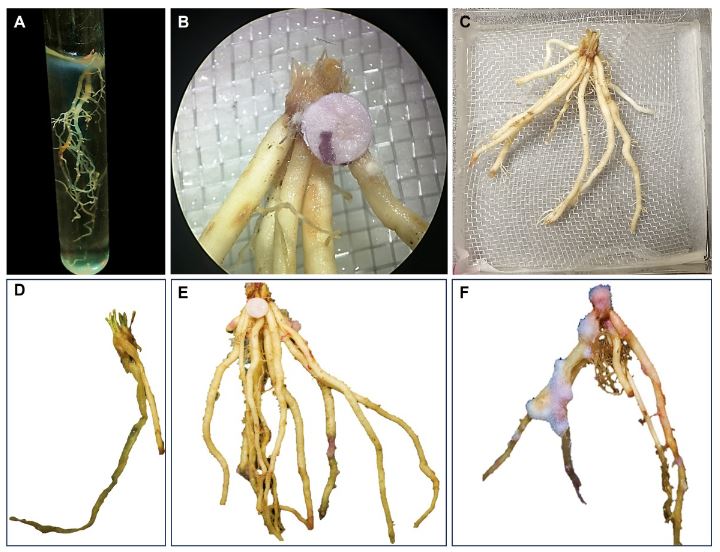
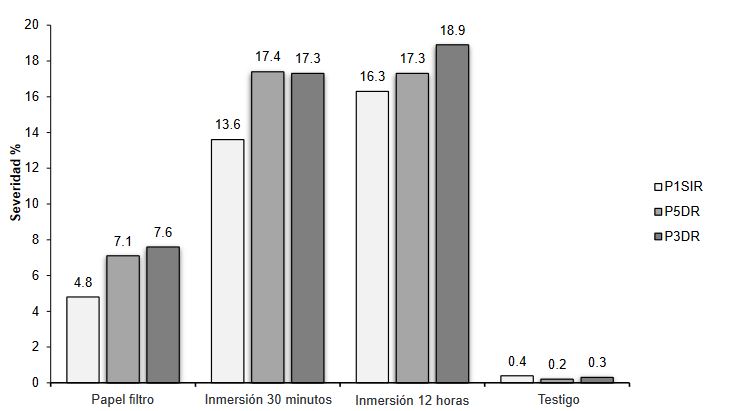
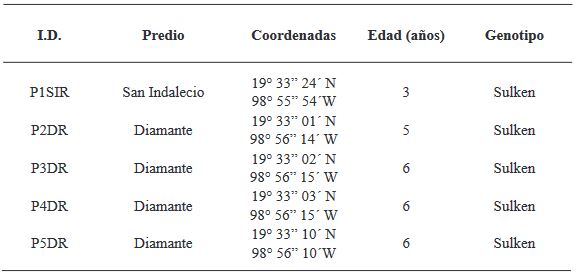
Materiales y Métodos. El muestreo se realizó en cinco parcelas productoras Atenco, Edo. de México. Se seleccionaron cinco aislados de Fusarium spp. (una por parcela) para realizar las pruebas de patogenicidad. Se seleccionaron tres aislados por sus características de colonización para las pruebas de severidad con diferentes métodos de inoculación: Inmersión por 12 h, inmersión por 30 min e inoculación por contacto de papel absorbente embebido en 1 mL de inóculo. Se emplearon con- centraciones de 1x106 de conidios mL-1. Se utilizaron 10 rizomas por tratamiento y 10 rizomas sin inocular. Para determinar la severidad, se analizaron fotografías (en GIMP®) del rizoma a los siete días después de la inoculación. Los aislados se identificaron molecularmente con ITS4/ITS5, EF688/EF1521 y TUBT1/BT2B.
Resultados. Se identificó morfológica y molecularmente a Fusarium prolifetatum en los tres aislados. El aislado P3DR fue el más severo (14.6%), seguido de P5DR (13.9%) y P1SIR (11.6%).
Conclusión. El método de inoculación más efectivo fue el de inmersión por 30 min. Se registraron en el Banco de Genes del NCBI con accesiones ON738484 (P3DR), ON973801 (P5DR) y ON738483 (P1SIR). Este es el primer reporte de F. prolifetatum en el Edo. de México.
-
O copiar el link