-
O copiar el link


Filtros de Contribuciones
Buscar Contribución:
por Jeny Michua Cedillo, Gustavo Mora Aguilera, Gerardo Acevedo Sánchez
Recibido: 30/7/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2406-7
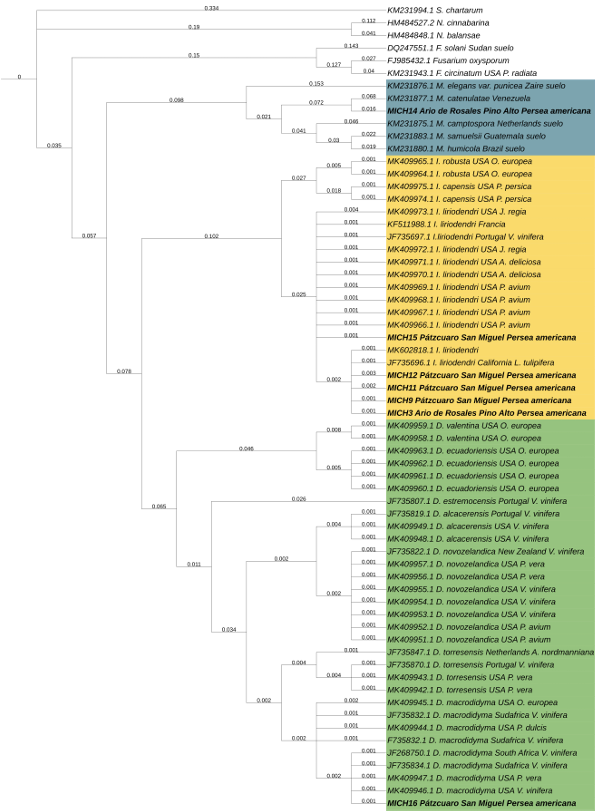
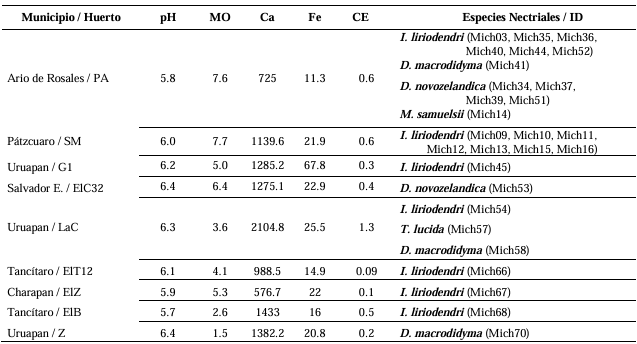
Resumen Antecedentes/Objetivo. Miembros de Nectriaceae se han detectado en Michoacán desde 2019. Sin embargo, su detección no ha tenido el enfoque etiológico primario por lo que se desconoce la identidad de especies en raíz, distribución geográfica y asociación con otras familias. El objetivo fue caracterizar biológica y molecularmente especies nectriales asociadas al aguacate (Persea americana).
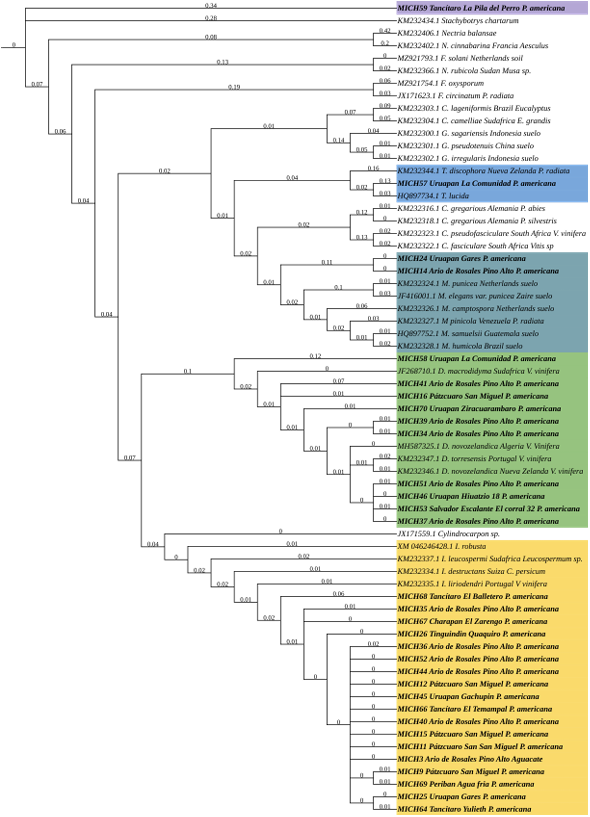
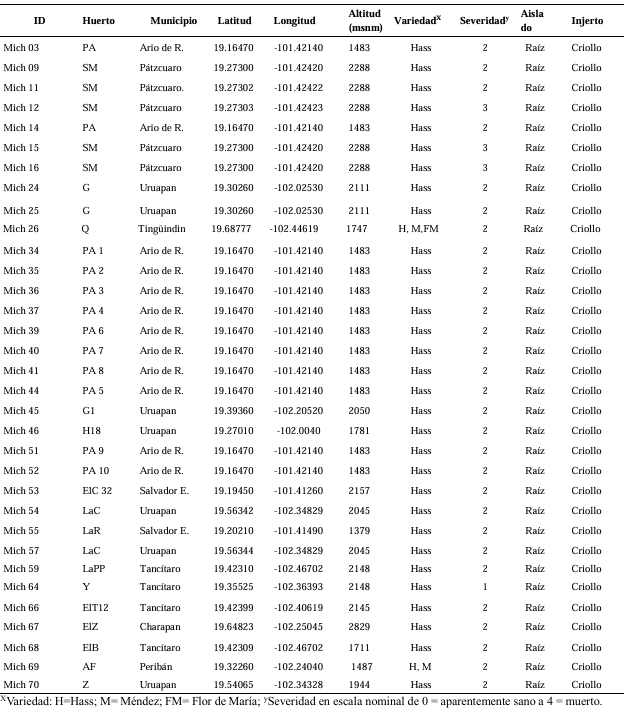
Materiales y Métodos. Se procesaron 70 muestras compuestas de raíces provenientes de árboles de aguacate con marchitez detectados en huertos comerciales de 13 municipios de Michoacán. Treinta aislados seleccionados con criterios epidemiológicos se cultivaron en extracto malta-agar, PDA y avena-agar para caracterización cultural y morfológica. A partir del ADN micelial se amplificaron los genes TEF 1-a y RPB2, las secuencias se limpiaron y alinearon con Seqassement y MAFFT, respectivamente. Se aplicaron algoritmos filogenéticos de inferencia Bayesiana y máxima parsimonia mediante PAUP4.0 y MrBayes3.2 complementándose con 66 y 65 secuencias del GenBank para TEF 1-a y RPB2, respectivamente. Cuatro especies de Hypocreales y S. chartarum se emplearon como especies externas.
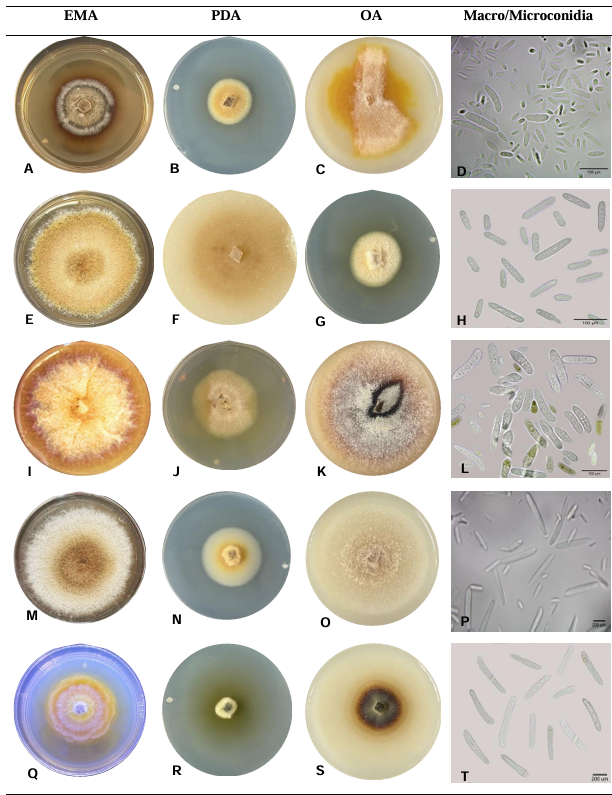
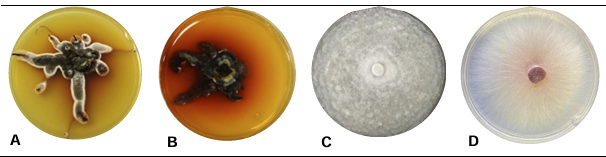
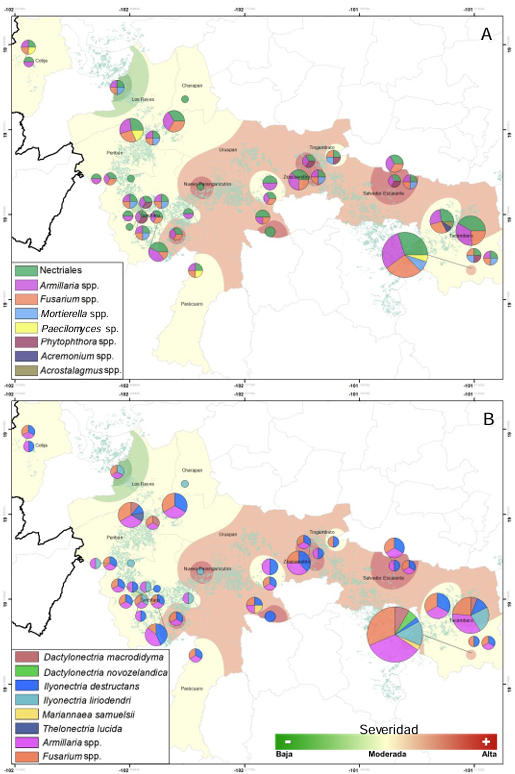
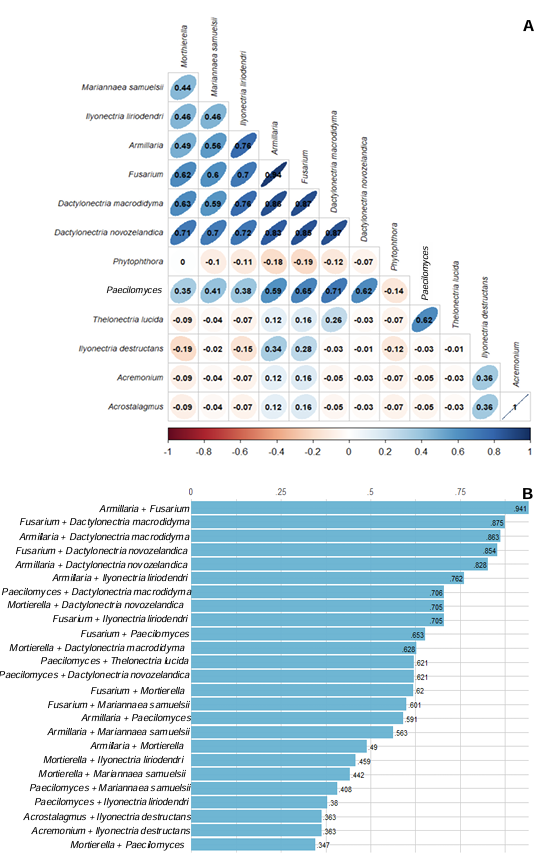
Resultados. Se obtuvieron cinco morfotipos de nectriales con crecimiento radial variable y coloración marrón. Se identificaron tres géneros y tres especies con TEF 1-a (>97% homología) y tres géneros y cinco especies con RPB2 (>97% homología) correspondientes a Ilyonectria (56% de los aislados), Dactylonectria (33 %), Mariannaea (6 %) y Thelonectria (3 %). En raíz, con niveles de daño variable, se observaron asociaciones significativas (p ≤ 0.05) entre especies nectriales con Armillaria (97.1 %), Fusarium (92.9 %), Paecilomyces (56.4 %) y Mortierella (47.3 %), no así con Phytophthora (r < - 0.07). Regionalmente, Ilyonectria liriodendry fue las más prevalente, preponderantemente asociada con Fusarium y/o Armillaria. El sur del municipio de Tacámbaro tuvo la mayor diversidad de especies nectriales y géneros fungosos en general.
Conclusión. Se postula la presencia regional de un síndrome de declinamiento en aguacate asociado a un complejo de hongos caracterizado por defoliación descendente, marchitez, reducción de crecimiento de fruto, decoloración y necrosis medular y cortical en raíz secundaria. Los nectriales Dactylonectria macrodidyma, D. novo-zelandica, Thelonectria lucida, Mariannaea samuelsii y I. liriodendry se asociaron con estos síntomas posiblemente con capacidad primaria de infección, pero frecuentemente en coinfección con Fusarium spp. y/o Armillaria spp. (r = 0.60 – 0.88, p ≤ 0.05). Otros hongos podrían estar asociados. Phytophthora podría tener limitada implicación en este síndrome. Este constituye el primer reporte de Ilyonectria, Dactylonectria, Mariannaea y Thelonectria asociados al aguacate (Persea americana) en México.
-
O copiar el link


por Ana Patricia Ibarra Valenzuela, Rosalba Troncoso Rojas, Alma Rosa Islas Rubio, Elizabeth Peralta, Herlinda Soto Valdez, Hayati Samsudin
Recibido: 09/4/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-4
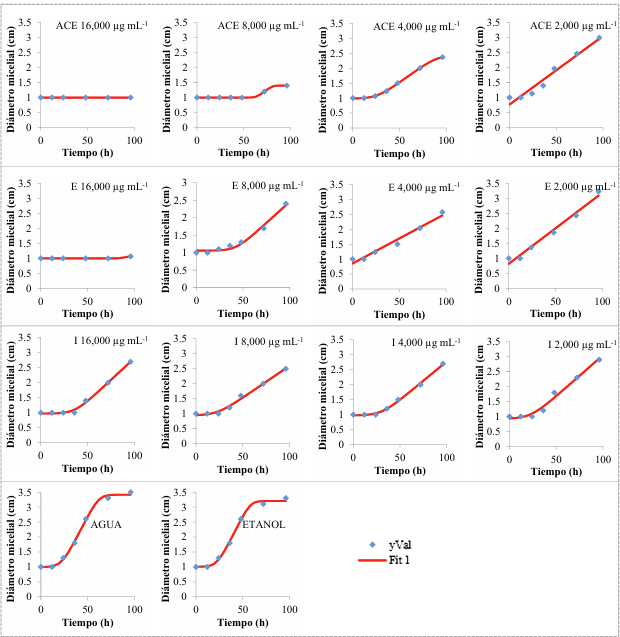
Resumen Antecedentes/Objetivo. Las tortillas de maíz, alimento básico en México, presentan una vida útil de 1-2 días a 25 °C debido al crecimiento fúngico. Una alternativa para extender la vida de anaquel de las tortillas es adicionar aceite esencial de clavo (AEC), sus componentes mayoritarios: eugenol (E), isoeugenol (I) y acetato de eugenilo (AE). El objetivo fue evaluar el efecto antifúngico del AEC sobre hongos identificados en tortillas de maíz.
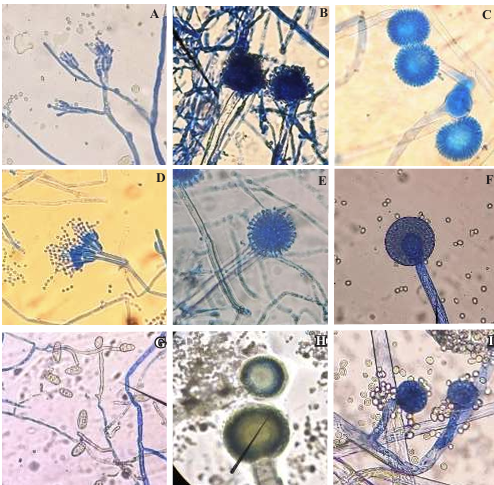
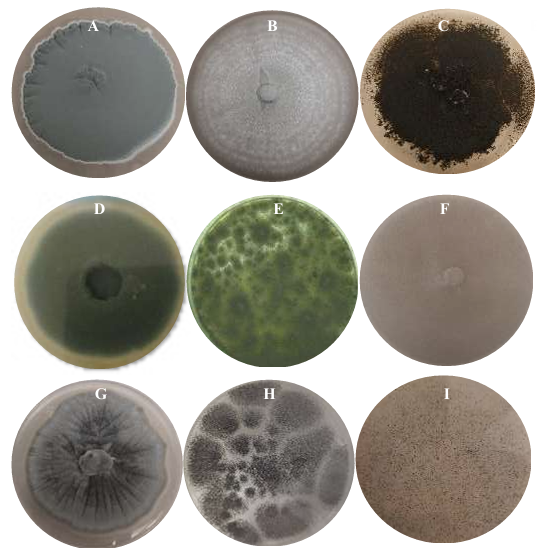
Materiales y Métodos. Se adquirieron muestras de un kg de tortillas de maíz de las capitales de cinco estados del país (Sonora, Nuevo León, Michoacán, Oaxaca y Yucatán). Los hongos se identificaron por su morfología y por biología molecular. Además, se les determinó la concentración mínima inhibitoria (CMI) frente a AEC. El efecto de E, I y AE sobre Aspergillus niger (previamente identificado) se evaluó con el modelo de Gompertz.
Resultados. Se obtuvieron dos aislados fúngicos de las tortillas de Nuevo León, Sonora, Yucatán y Michoacán y un aislado de Oaxaca, mismos que se identificaron por biología molecular: Aspergillus longivesica y Curvularia spicifera de Nuevo León; Aspergillus niger y Penicillium brevicompactum de Sonora; Aspergillus sp. de Oaxaca; Mucor sp. y Aspergillus flavus de Yucatán; Penicillium herquei, y Curvularia racemosus de Michoacán. Las CMIs fueron 200, 400, 800, 400, 800, 400, 800, 800 y 400 µg mL-1, respectivamente. AEC, E e I a 800 µg mL-1 retardaron la fase exponencial de crecimiento de Aspergillus niger, mientras que AE no mostró efecto.
Conclusión. El AEC podría ser una alternativa natural para prolongar la vida útil de tortillas de maíz.
-
O copiar el link


por Augusto Gil Ceballos Ceballos, Yisa María Ochoa Fuentes, Ernesto Cerna Chávez, Arely Cano García
Recibido: 18/4/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-5
Resumen Antecedentes/Objetivo. Los extractos de semillas de Canavalia ensiformis han demostrado efectos antiparasitarios y repelentes contra plagas. El objetivo fue evaluar la efectividad del extracto combinado con nanopartículas (NP's) de dióxido de silicio contra los huevos de Meloidogyne incognita.
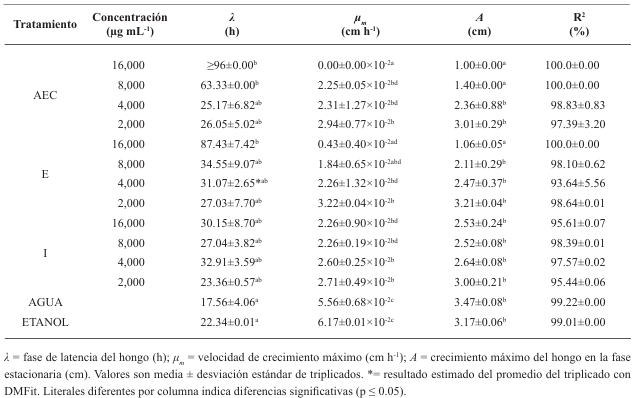
Materiales y Métodos. Se llevaron a cabo experimentos in vitro para evaluar los efectos de los extractos de semillas de C. ensiformis, tanto por sí solos como combinados con NP's de dióxido de silicio, en la eclosión de juveniles de M. incognita. Se utilizaron 150 huevos y se aplicaron concentraciones de 0, 2, 4, 6, 8 y 10 % del extracto. Además, se evaluaron concentraciones del extracto al 0, 1.5, 2.0, 2.5 y 3.0 %, cada una combinada con concentraciones de NP's al 0.06, 0.08, 0.10, 0.12 y 0.14 %.
Resultados. Ninguno de los tratamientos logró evitar la eclosión más del 30 % de juveniles. La modificación de la técnica de obtención del extracto de semillas de C. ensiformis puede causar un efecto ovicida complementario; sin embargo, al aumentar las concentraciones del extracto, puede propiciar la proliferación de hongos saprofitos y otros microorganismos.
Conclusión. Los tratamientos no mostraron efectos ovicidas significativos arriba del 40 %.
-
O copiar el link
Estimación de pérdidas provocadas por Potato virus Y en el cultivo de papa en Coahuila
por Joel De Santiago Meza, Gustavo Alberto Frías Treviño, Luis Alberto Aguirre Uribe, Alberto Flores Olivas
Recibido: 05/4/2024 – Publicado: 30/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-2
Resumen Antecedentes/Objetivo. El objetivo fue evaluar experimentalmente las pérdidas ocasionadas por PVY (Potato Virus Y) en el cultivo de papa variedad Fianna y, por consiguiente, estimar las pérdidas ocasionadas por este virus en la zona productora de papa de Coahuila.
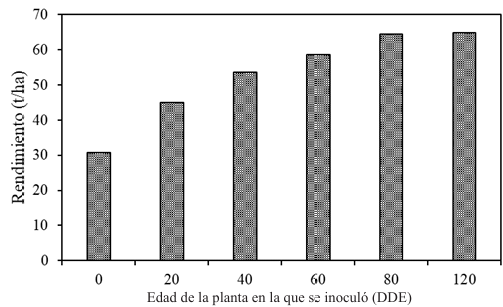
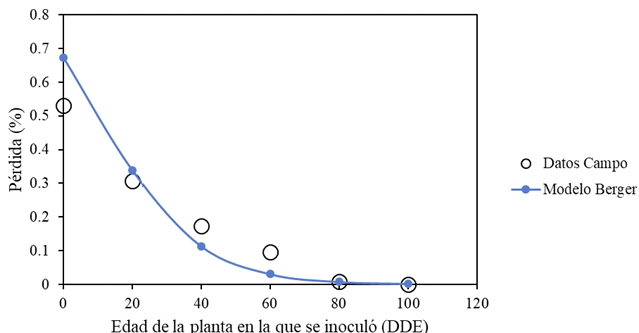
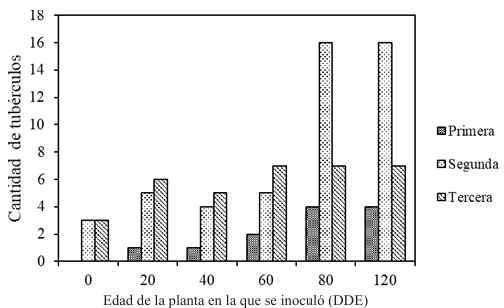
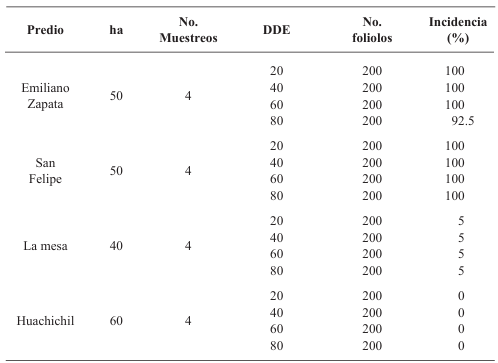
Materiales y Métodos. Surcos de una parcela experimental sembrados con plántula y semilla-tubérculo de papa, se inocularon mecánicamente con PVY a los 20, 40, 60 y 80 días después de la emergencia. Se cosecharon los tubérculos producidos y se evaluaron las pérdidas en cada tratamiento. Adicionalmente, en cuatro predios comerciales de papa en este mismo estado, se tomaron muestras de foliolos a los 20, 40, 60 y 80 días después de la emergencia, y se evaluó mediante pruebas de ELISA el porcentaje de plantas infectadas con PVY. Los datos de pérdidas de la parcela experimental y los datos de incidencia de los predios se utilizaron para elaborar un modelo estadístico para estimar las pérdidas causadas por PVY en la región de Coahuila.
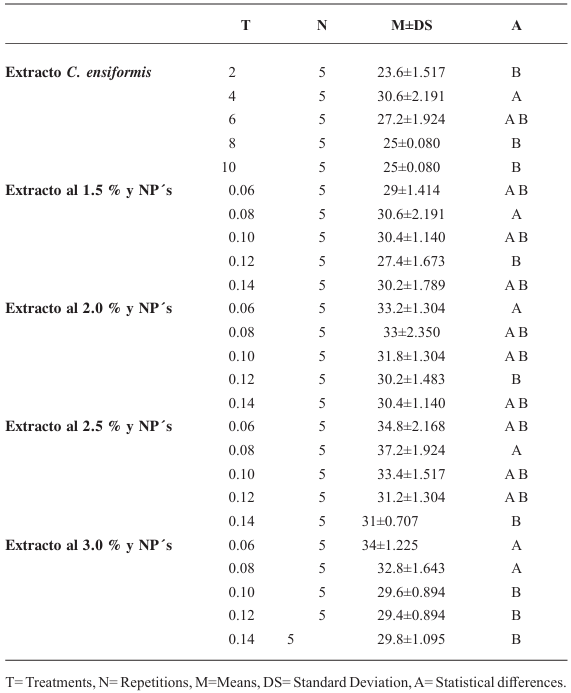
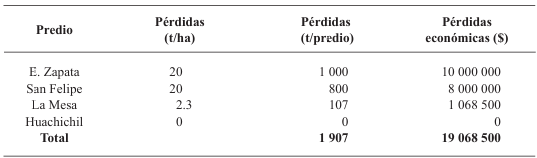
Resultados. Las pérdidas en el rendimiento por PVY en la parcela experimental fueron de 9.4% a 53%. El porcentaje de incidencia de plantas infectadas en los predios comerciales varió de 0% a 100%. El modelo que mejor se ajustó a los datos obtenidos fue el de Berger . Las pérdidas estimadas en la zona productora de papa de Coahuila en el ciclo 2022 fueron de 18 %, equivalente a $19, 068, 500.
Conclusión. Esta información resalta la importancia de utilizar semilla certificada libre de PVY y de proteger el cultivo desde la emergencia hasta los 60 DDE.
-
O copiar el link


por Alejandra Chacón López, José Luis Hernández Flores, Efigenia Montalvo González, Selene Aguilera Aguirre
Recibido: 20/8/2024 – Publicado: 30/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2308-2
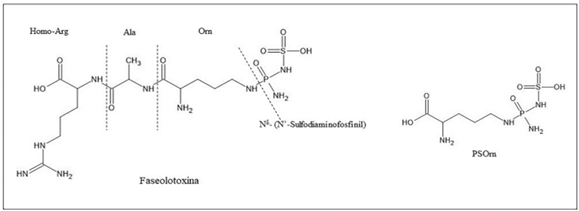
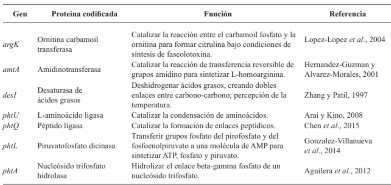
Resumen Antecedentes/Objetivo. La faseolotoxina es producida por uno de los fitopatógenos más importantes y estudiados en el área agrícola: Pseudomonas syringae pv. phaseolicola. Esta bacteria causa el tizón de halo, una enfermedad que devasta al cultivo del frijol. El éxito de P. syringae pv. phaseolicola está relacionado con su información genética que le permite sintetizar metabolitos deletéreos para su hospedero, como la faseolotoxina. El objetivo de la presente investigación fue analizar la base molecular del mecanismo de acción, la inmunidad, la genética involucrada en la biosíntesis de la faseolotoxina, las estrategias de diagnóstico molecular y las técnicas moleculares desarrolladas en México, para llevar a cabo el manejo del tizón de halo del frijol.
Materiales y Métodos. Se realizó la búsqueda y el análisis de la información científica más relevante respecto a la biosíntesis de faseolotoxina y los estudios moleculares de los factores de patogenicidad y virulencia de P. syringae pv. phaseolicola que han contribuido al desarrollo de estrategias moleculares enfocadas en el diagnóstico y manejo del tizón de halo en frijol.
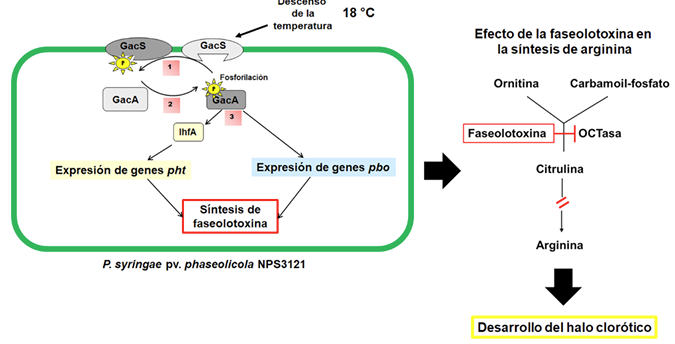
Resultados. P. syringae pv. phaseolicola produce faseolotoxina, que es la responsable de la formación del halo clorótico característico del tizón de halo, esta toxina es un inhibidor de la OCTasa, una enzima que participa en la ruta de síntesis de arginina en frijol. Las regiones cromosómicas Pht y Pbo contienen genes involucrados en la síntesis e inmunidad de la faseolotoxina, y la expresión de estos genes está regulada por el sistema GacS/GacA y la temperatura. La identificación de genes involucrados en la síntesis de factores de patogenicidad y virulencia, como la faseolotoxina, ha permitido el desarrollo de estrategias de diagnóstico y manejo de la enfermedad basadas en la amplificación de ADN y el uso de marcadores moleculares que facilitan la identificación de cultivares de frijol resistentes al patógeno.
Conclusión. Los estudios moleculares han contribuido al entendimiento de cómo el patovar phaseolicola produce faseolotoxina. Esta información ha sido esencial para entender cómo las bacterias han evolucionado de variantes no patogénicas a patogénicas; además, proporcionan información que permite desarrollar nuevas estrategias para un diagnóstico oportuno y contribuyen en las estrategias para el manejo del tizón de halo.
-
O copiar el link




por Conrado Parraguirre Lezama, Omar Romero Arenas, Alba Cruz Coronel, Amparo Mauricio Gutiérrez, Carlos A Contreras Pare, Antonio Rivera Tapia
Recibido: 25/2/2024 – Publicado: 27/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2402-7
Resumen Objetivo/Antecedentes. La transición hacia prácticas agrícolas responsables es fundamental para promover la salud de los agroecosistemas y garantizar la seguridad alimentaria. Impulsar investigaciones integrales que combinen métodos químicos y biológicos representa un avance significativo en el manejo de fitopatógenos, es decir, esta aproximación novedosa se basa en la premisa de que la acción conjunta entre fungicidas y un agente antagónico como Trichoderma spp., pueden ofrecer una protección robusta en comparación con enfoques individuales. El objetivo del estudio es investigar la resistencia y compatibilidad in vitro de cuatro especies de Trichoderma frente a tres fungicidas ampliamente utilizados en México.
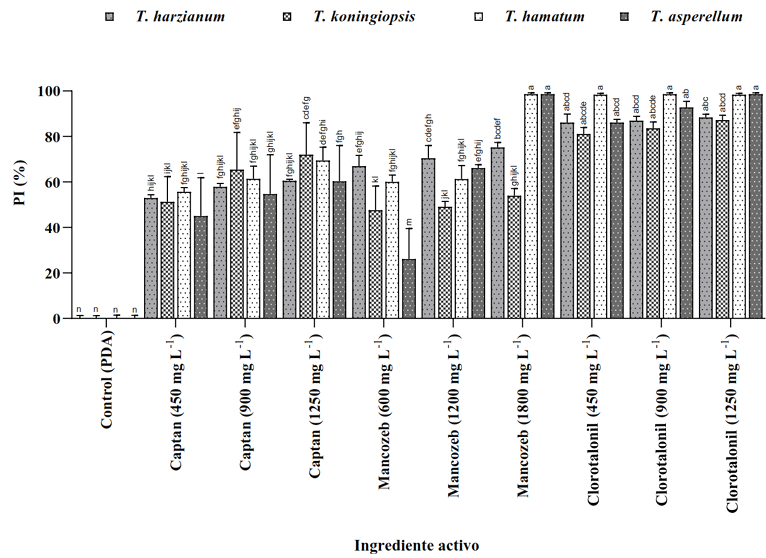
Materiales y Métodos. Se empleó la técnica de intoxicación controlada en medio PDA bajo condiciones controladas con tres concentraciones (450, 900 y 1350 mg L−1), pare el caso de los ingredientes activos Captan y Clorotalonil, para Mancozeb se utilizaron 600, 1200 y 1800 mg L−1. La compatibilidad se determinó en relación con el grupo control utilizando el software estadístico SPSS Statistics versión 26 para el entorno operativo Windows.
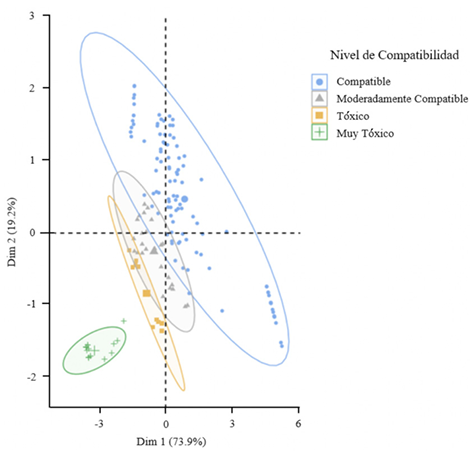
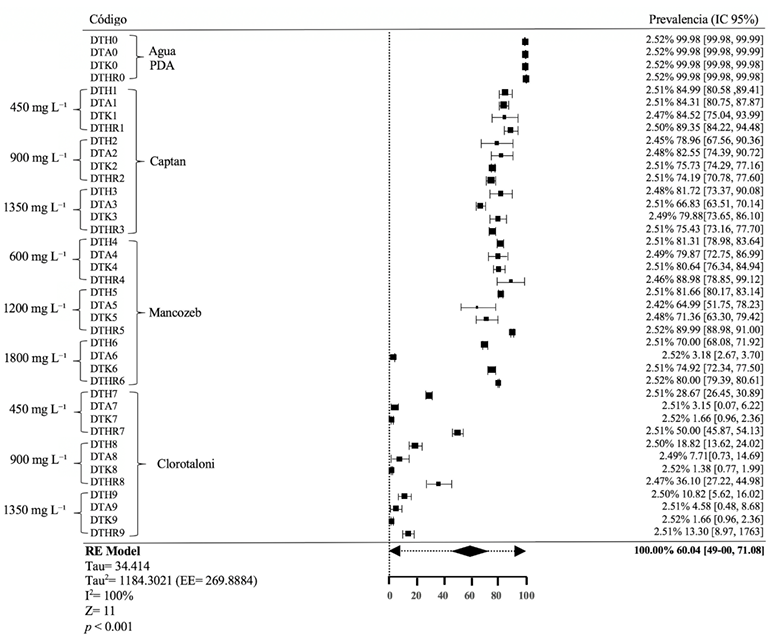
Resultados. El estudio reveló que las cepas de T. harzianum, T. hamatum, T. koningiopsis y T. asperellum exhibieron una compatibilidad global del 60.04% para los ingredientes activos evaluados, siendo el fungicida Captán 50® el que demostró el mayor porcentaje de compatibilidad (79.87%) en las concentraciones de 450, 900 y 1350 mg L–1. T. harzianum mostró mayor tolerancia al ingrediente activo Clorotalonil en la concentración de 450 mg L⁻¹, sin embargo, a concentraciones más altas demostró mayor toxicidad, siendo T. koningiopsis la que exhibió la menor resistencia en sus tres concentraciones evaluadas.
Conclusión. Los tratamientos con diferentes concentraciones de los fungicidas Captan, Mancozeb y Clorotalonil evidenciaron una marcada variabilidad en términos de prevalencia y toxicidad hacia las especies evaluadas de Trichoderma in vitro. Este enfoque permite diseñar estrategias de manejo integrado minimizando la dependencia de productos químicos y promoviendo la compatibilidad entre agentes biológicos y fungicidas.
-
O copiar el link
Etiología de la pudrición marrón tostado en fresa (Fragaria x ananassa) en el Estado de México
por Hugo Velasco Montaño, Victoria Ayala Escobar, Daniel Téliz Ortiz, Nadia Landero Valenzuela, Santos Gerardo Leyva Mir
Recibido: 07/6/2024 – Publicado: 27/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2406-3
Resumen Antecedentes/Objetivo. En cultivo de fresa establecidos en invernadero en Montecillo, Texcoco, Estado de México, en 2022 se observó manchas foliares color marrón tostado y pudrición de frutos con lesiones hundidas asimétricas, que se extendían y adquirían un color marrón. El objetivo del presente trabajo fue identificar el agente causal de la pudrición marrón tostado en frutos y plantas fresa.
Materiales y Métodos. Se colectaron frutos y hojas sintomáticos, de los cuales se obtuvieron aislados fúngicos para realizar las pruebas de patogenicidad en plantas y frutos, en plantas mediante dos métodos de inoculación: aspersión vía foliar y vía raíz; en frutos mediante inmersión. Se emplearon concentraciones de 2×106 conidios mL−1. Se amplificó y secuenció la región ITS del rDNA mediante PCR con los iniciadores universales ITS1-ITS4.
Resultados. Se identificó morfológica y molecularmente a Pilidium concavum como el agente causal de la mancha y pudrición marrón tostado en fresa. Resultó patogénica en frutos de fresa cv. Aromas y en plantas menores de dos meses de edad. Mostró variación en virulencia, en plantas afectadas varió de 40 a 50%, en frutos alcanzó el 100%.
Conclusión. El resultado determina que Pilidium concavum es un patógeno que produce mancha foliar marrón tostado y pudrición marrón tostado en frutos de fresa. Permite nuevas líneas de investigación relacionados con el impacto de la enfermedad en la producción, rendimiento y calidad de fresas en México. Esta investigación es el primer reporte de Pilidium concavum como patógeno de fresa en el Estado de México.
-
O copiar el link


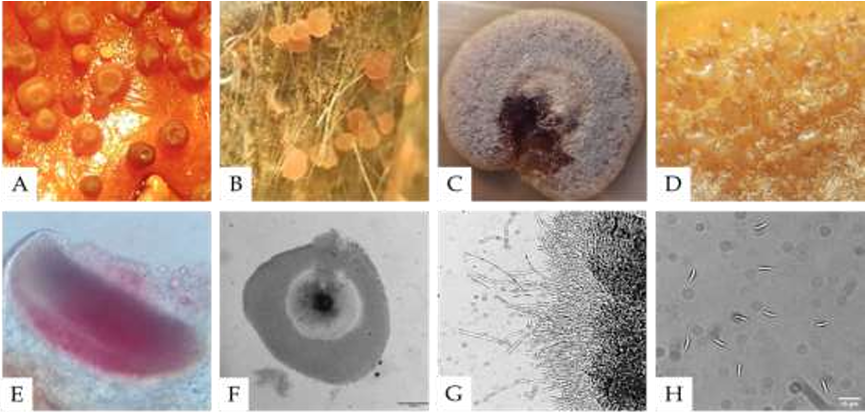




Diversidad y taxonomía de Fusarium solani aislado de plantas marchitas de Agave tequilana var azul
por Viviana Montaño Becerrra, Norma Alejandra Mancilla Margalli, Cristina Chávez Sánchez, Martin Eduardo Avila Miranda
Recibido: 30/11/2023 – Publicado: 25/10/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-5
Resumen Antecedentes/Objetivo. El objetivo del trabajo fue identificar 24 cepas de F. solani aisladas de agave con marchitez, respecto a las nuevas especies filogenéticas; determinar su semejanza molecular con f. spp. de F. solani; determinar su diversidad genética y su capacidad patogénica en agave, frijol (Phaseolus vulgaris) y maíz (Zea mays).
Materiales y Métodos. Secuencias del fragmento ITS1-5.8S-ITS2 de 24 aislados de agave y de las f. spp. de F. solani, se compararon con el GenBank y FUSAROIDID. Secuencias amplificadas del 18S rRNA, se alinearon con secuencias reportadas de F. solani f. spp. phaseoli y batatas, definiendo presencia de intrones. Se determinó diversidad genética con el marcador de DNA RepPCR. Cepas representativas se confrontaron con plántulas de agave, frijol y maíz, evaluando su patogenicidad como severidad de pudrición radicular.
Resultados. Aislados morfológicamente identificados como F. solani, el GenBank los ubicó como F. solani o incluidos en el FSSC, tres cepas se identificaron como Xenoacremonium sp. FUSAROID-ID definió que las secuencias de F. solani eran altamente similares a las de Neocosmospora martii, N. pseudoradicicola, N. solani y N. falciformis. Las secuencias ITS1-5.8S-ITS2 y ausencia de intrones en su SSU, indicó que ninguno es F. solani f. sp. phaseoli. Aislados obtenidos de agave fueron patogénicos a A. tequilana y a un cv de maíz criollo, pero no a maíces con resistencia a Fusarium. Ningún aislado de agave fue patogénico a frijol.
Conclusiones. Cuatro especies filogenéticas del FSSC provocan pudrición radicular en agave; aislados de F. solani de agave no afectaron a maíces resistentes a Fusarium. Es seguro intercalar frijol en el cultivo de agave.
-
O copiar el link

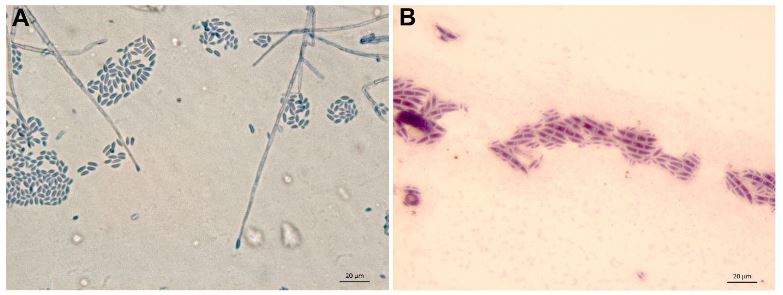
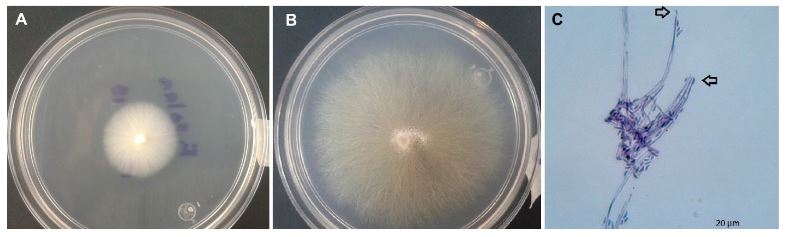
Tobamovirus fructirugosum una enfermedad emergente: revisión y situación actual en México
por Ubilfrido Vásquez Gutiérrez, Juan Carlos Delgado Ortiz, Gustavo Alberto Frías Treviño, Luis Alberto Aguirre Uribe, Alberto Flores Olivas
Recibido: 28/1/2024 – Publicado: 15/10/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2401-7
Resumen Antecedentes/Objetivo. La especie Tobamovirus fructirugosum (ToBRFV) es considerado una plaga cuarentenaria a nivel mundial que limita la producción de Solanum lycopersicum y Capsicum annum, actualmente presente en tres países del continente americano. El objetivo de este trabajo fue profundizar en la variabilidad genética del ToBRFV con respecto a los diversos aislados, la caracterización físico-molecular y sintomática, los métodos tradicionales y más actuales implementadas para el diagnóstico, rango de hospedantes reservorios del virus, y la epidemiología. Resultados. ToBRFV se generó de una mutación resultado de la recombinación genética con TMV, considerado principal progenitor y ToMMV progenitor secun¬dario. Análisis filogenéticos reportan la existencia de cinco clados con respecto a la diversidad genética del ToBRFV. Los primeros cebadores para la detección se diseñaron en 2015 que codifican proteínas de replicación, movimiento y cápside. Los métodos serológicos pueden ser utilizados para un diagnóstico preventivo, mientras que las moleculares y NGS pueden confirmar la infección por el virus aún en bajas concentraciones en la planta. Se reportan 16 familias de malezas y cultivos hospedantes, registrados en 47 países. Para lograr una estrategia efectiva, es necesario disminuir las fuentes de inóculo, desarrollar compuestos inhibidores de la transmisión mecánica y el desarrollo de genotipos tolerantes. Conclusión. ToBRFV está distribuido a nivel nacional, y representa un riesgo fitosanitario para México; el análisis exhaustivo del estudio de técnicas de diagnóstico, rango de hospedantes, diseminación, epidemiología y estrategias de control, contribuye al conocimiento del ToBRFV.
-
O copiar el link