

-
O copiar el link


Filtros de Contribuciones
Buscar Contribución:
PorOscar Guillermo Rebolledo Prudencio, Wilberth Chan Cupul*, Guadalupe Moreno Zúñiga, Carlos Adrián Cruz Jiménez, Juan Carlos Sánchez Rangel
Recibido: 15/11/2024 – Publicado: 21/2/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-09
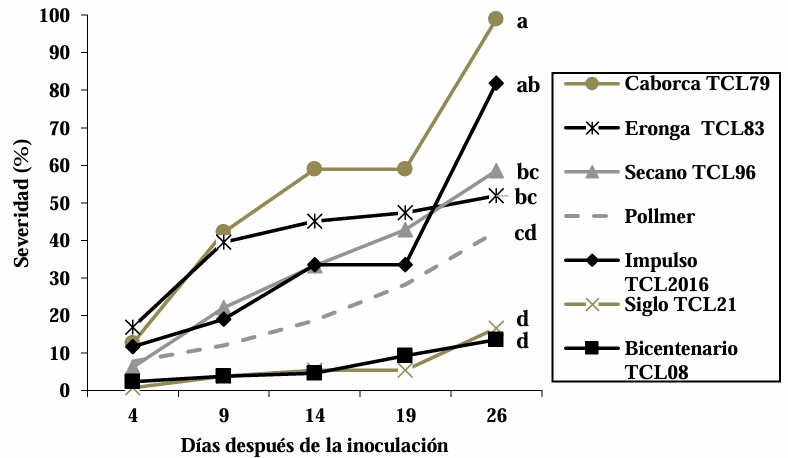
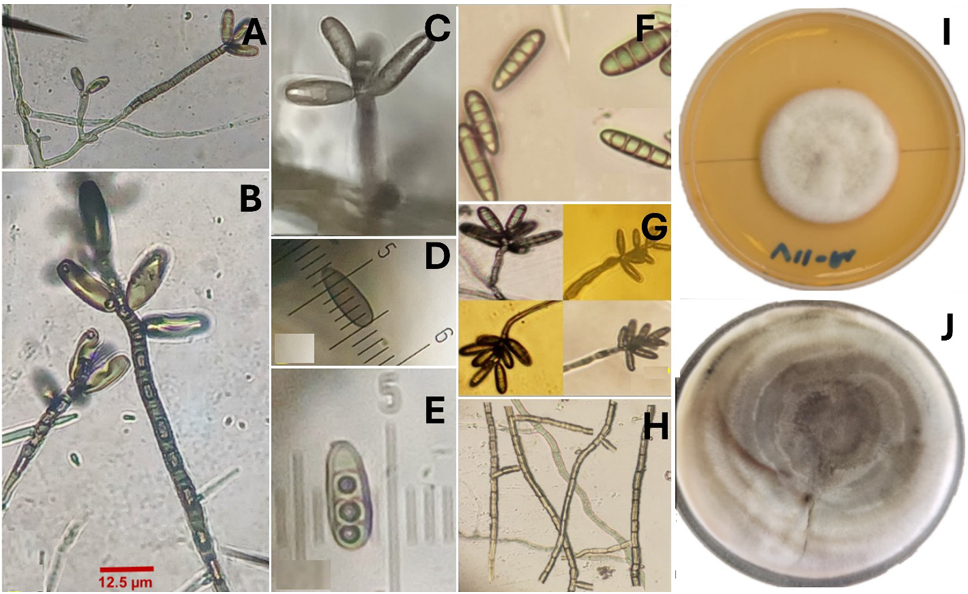
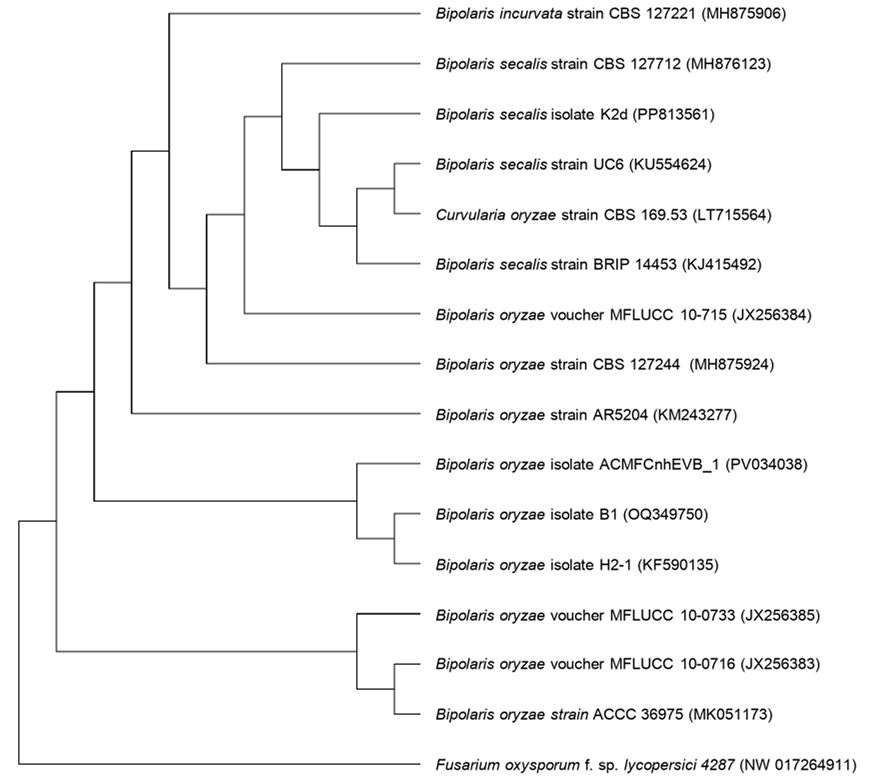
Resumen Antecedentes/Objetivo. En Tecomán, Colima, México, se detectó una mancha foliar (MF) con incidencia del 92.0% en Cocos nucifera híbrido “Enano Verde de Brasil” (EVB). El objetivo fue caracterizar morfológica, molecular y bioquímicamente al hongo asociado a la MF en palma de coco EVB y evaluar su susceptibilidad a fungicidas biológicos comerciales.
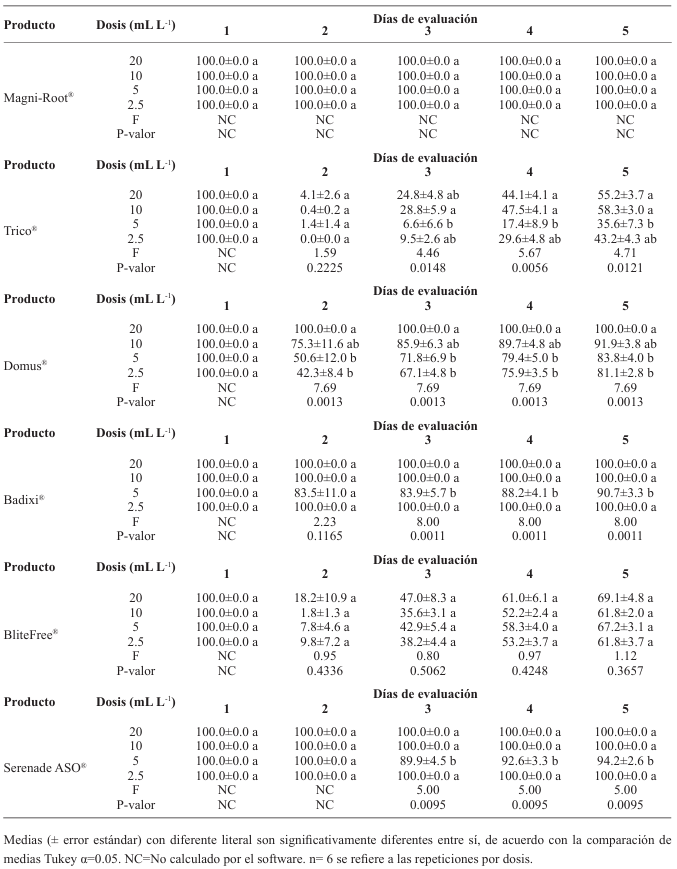
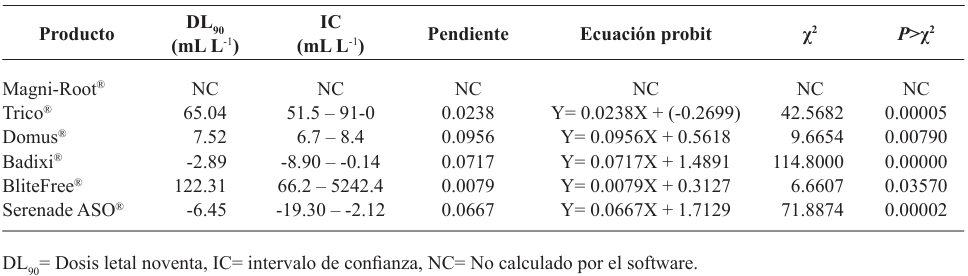
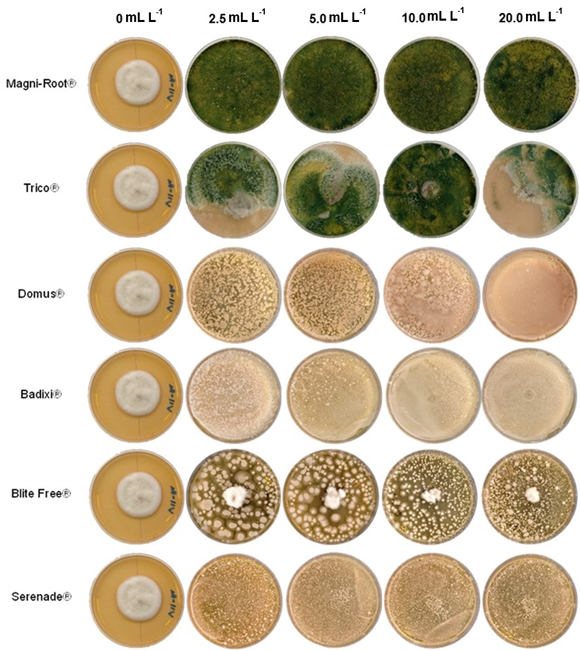
Materiales y métodos. El aislado se caracterizó morfológica y molecularmente. Se evaluó su crecimiento, esporulación y producción de lacasas en diferentes medios de cultivo. Se determinó la inhibición micelial in vitro y dosis letal media (DL50) de fungicidas biológicos comerciales a base de hongos antagonistas (Trichoderma harzianum y T. viride), bacterias (Bacillus subtilis y B. amyloliquefaciens) y actinobacterias (Streptomyces lydicus y S. jofer).
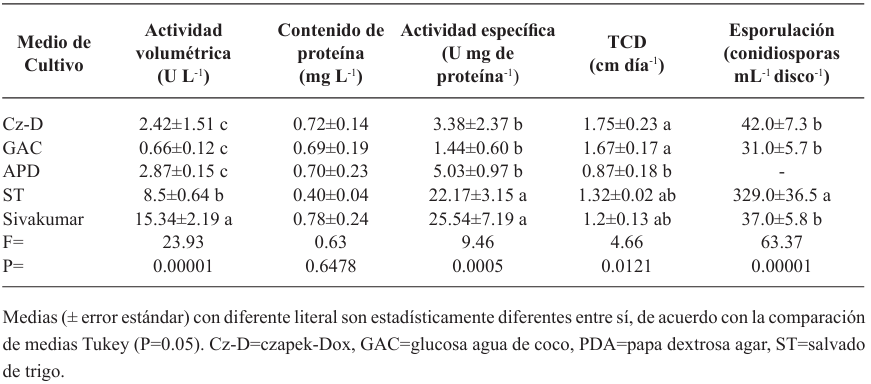
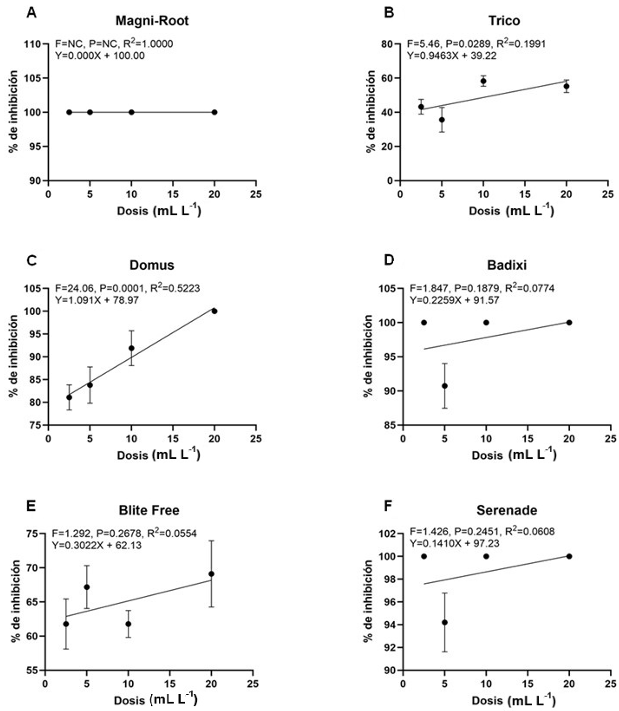
Resultados. Bipolaris oryzae fue el agente asociado a la MF, produjo 25.54 y 22.17 U mg de proteína-1 de actividad lacasa en los medios Sivakumar y salvado de trigo (ST). El medio ST permitió la mayor esporulación. T. harzianum inhibió al 100% a B. oryzae en las cuatro dosis evaluadas. B. subtilis y B. amyloliquefaciens inhibieron al 100% a B. oryzae en la dosis más alta evaluada (20 mL L-1).
Conclusión. Bipolaris oryzae es el agente asociado a la MF, produjo la mayor actividad lacasa en Sivakumar y ST. La mayor esporulación y crecimiento diario fue en ST. T. harzianum destacó sobre T. viride al inhibir en 100% el crecimiento de B. oryzae. Bacillus subtilis, S. lydicus y B. amyloliquefaciens fueron más efectivas contra B. oryzae comparado con S. jofer.
-
O copiar el link
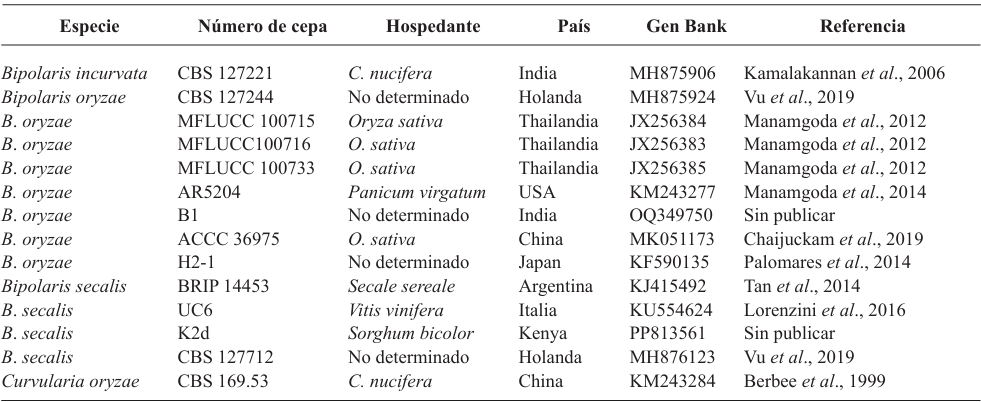

PorMaría del Sol Cuellar Espejel, Evangelina Esmeralda Quiñones Aguilar, Gabriel Rincón Enríquez*, Rodolfo Hernández Gutiérrez, Juan Carlos Mateos Díaz, Sergio David Valerio Landa
Recibido: 15/11/2024 – Publicado: 13/2/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-25
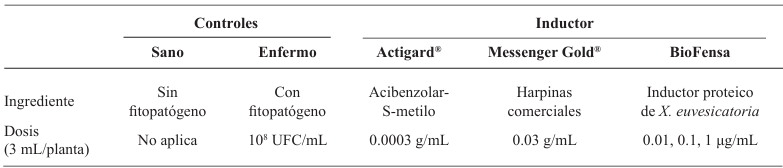
Resumen Antecedentes/Objetivo. Las enfermedades como la peca bacteriana en jitoma te (Solanum lycopersicum) y la mancha bacteriana en chile (Capsicum annuum) causan graves pérdidas económicas globales. Una alternativa sustentable para su control es el uso de inductores proteicos (proteínas Harpin=Hrp) que activan la respuesta de defensa de las plantas al ser reconocidas por el sistema de inmunidad vegetal, induciendo mecanismos de defensa en contra de patógenos. El objetivo de esta investigación fue evaluar la efectividad biológica y la dosis óptima de apli cación del inductor biológico BioFensa (a base de proteínas Hrp), producido en planta piloto para controlar estas enfermedades.
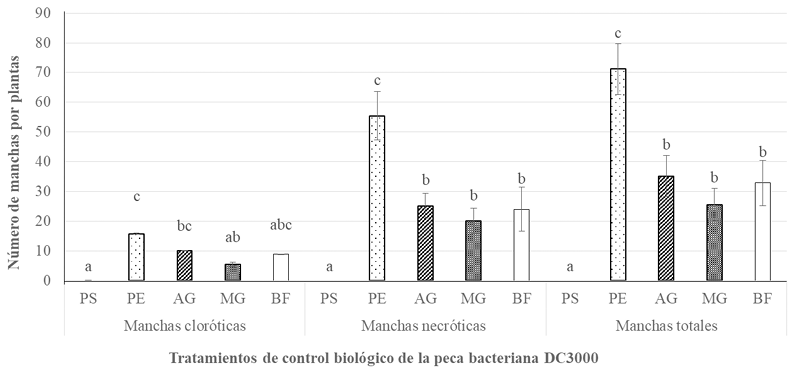
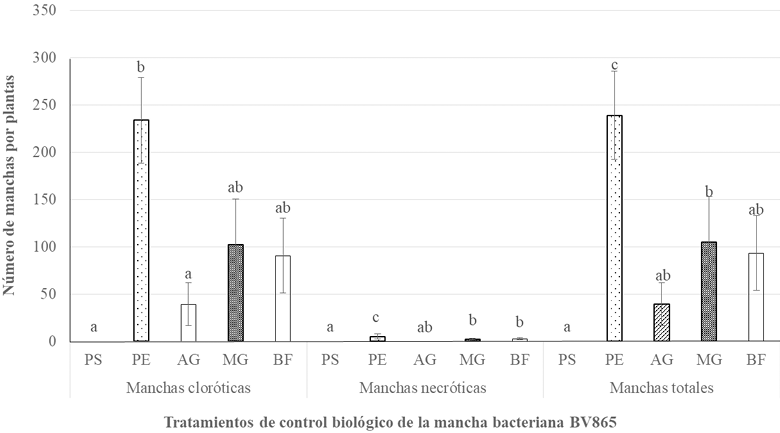
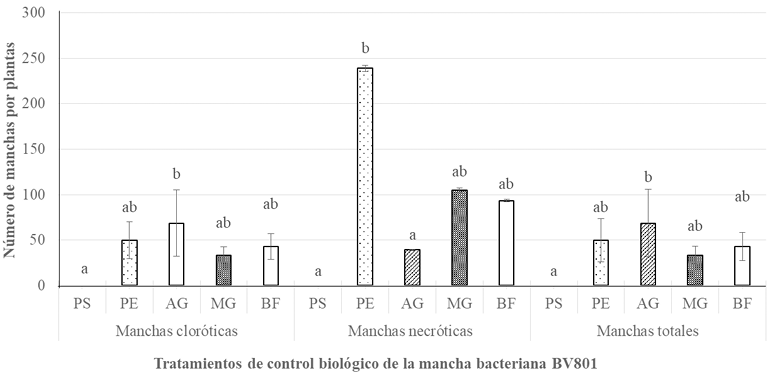
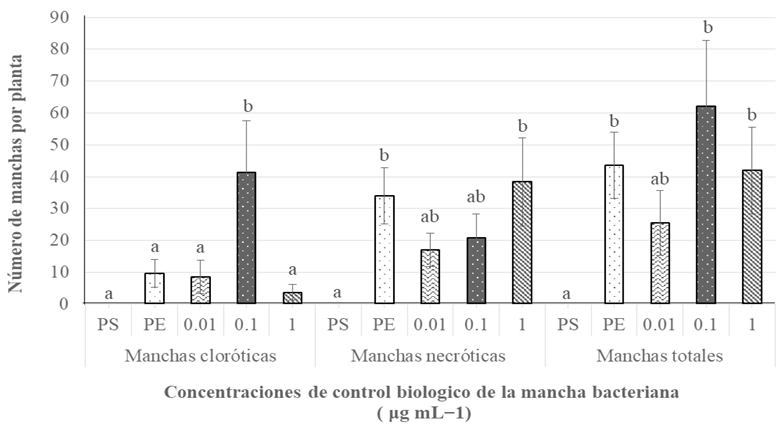
Materiales y Métodos. Se realizaron tres experimentos en invernadero para eva luar la efectividad biológica de Biofensa (1 μg mL-1); para ello se probó el inductor proteico para el control de la mancha bacteriana (X. euvesicatoria cepa BV865 [1] y BV801 [2]), así como para la peca bacteriana (P. syringae pv. tomato, cepa DC3000 [3]). Cada experimento incluyo 5 tratamientos y 11 repeticiones. Además, se realizó un experimento para determinar la dosis óptima de BioFensa (0.01, 0.1 y 1.0 μg mL-1) contra X. euvesicatoria cepa BV801, con 7 tratamientos y 8 repeti ciones [4]. En los cuatro experimentos en total, las plantas fueron asperjadas con BioFensa (3 mL por planta) 24 h antes de la infección y se evaluaron los síntomas después de 30 días contando manchas en el tejido foliar.
Resultados. BioFensa fue efectivo en reducir significativamente el daño en plan tas de chile y tomate (LSD, p≤0.05). A una concentración alta (1 μg mL-1) logró prevenir la aparición de manchas en plantas de jitomate en un 53%, mientras que para plantas de chile contra la cepa BV865 previno en un 60%. Por otro lado, para para plantas de chile contra la cepa BV801, a concentraciones bajas (0.01 y 0.1 μg mL-1), los síntomas se redujeron significativamente entre un 38-41%, mientras que a una concentración más alta (1 μg mL-1) este efecto no se mantuvo, sugiriendo un límite en la percepción de inductores por las plantas.
Conclusión. Los resultados sugieren que BioFensa tiene el potencial de ser una alternativa efectiva para controlar enfermedades en cultivos hortícolas como jito mate y chile.
-
O copiar el link
PorWilberth Chan Cupul*, Osvaldo Villegas Guerrero, Juan C. Sánchez Rangel, Gilberto Manzo Sánchez, Marco T. Buenrostro Nava
Recibido: 15/11/2024 – Publicado: 13/2/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-05
Resumen Antecedentes/objetivo. La Sigatoka negra (SN) es una de las principales fitopatologías que reduce la producción de banano en México. Desarrollar productos biológicos a base de antagonistas es una actividad preponderante de estudio. Se evaluó la producción de conidiosporas de Trichoderma spp. en fermentación sólida sobre granos de arroz y maíz, y se estudió el efecto in situ de conidiosporas y Streptomyces sp. en la epidemiologia de la SN en banano cv. Gran enano.
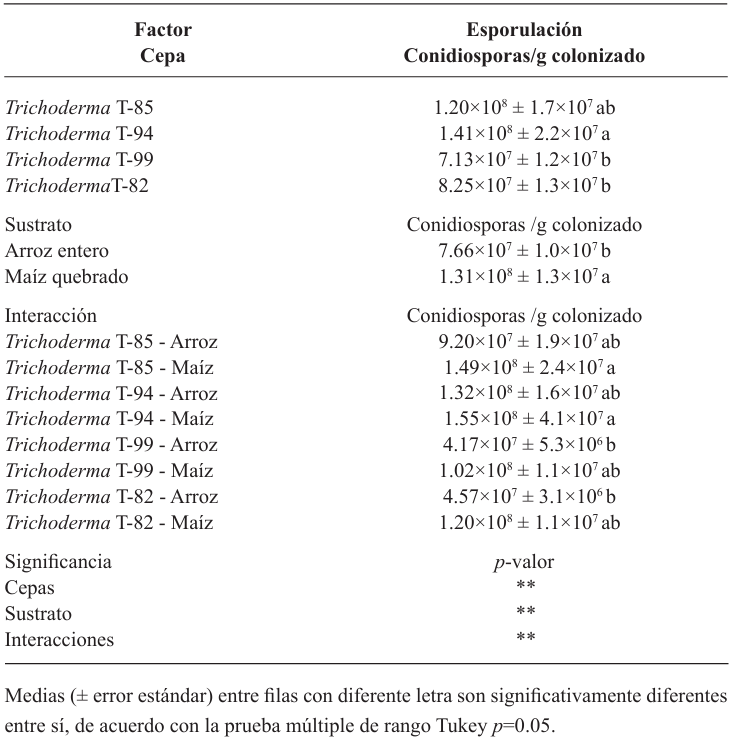
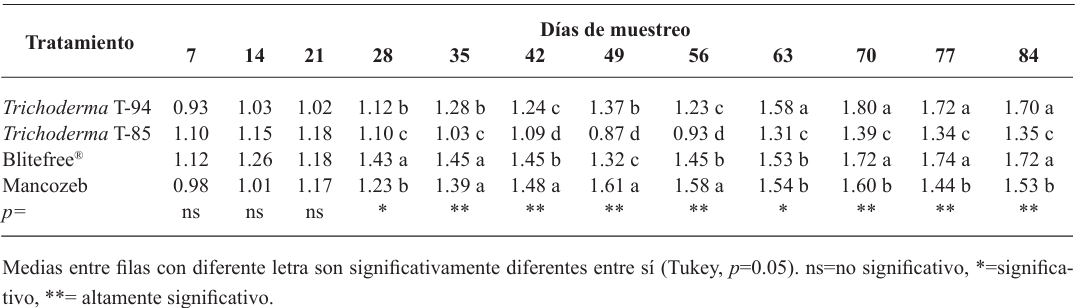
Materiales y métodos. En fermentación sólida se evaluó el rendimiento de cuatro cepas de Trichoderma spp. (T-82, T-85, T-94 y T-99) en arroz entero (AE) y maíz quebrado (MQ); se empleó un diseño factorial A×B (A=cepas y B=sustrato). Las dos cepas con mejor rendimiento (T-99 y T-85) y un producto a base de Streptomyces spp. se aplicaron en campo para evaluar la epidemiología de la SN a través de la severidad, promedio ponderado de infección (PPI) y área bajo la curva del progreso de la enfermedad (ABCPE), a través de un diseño de bloques al azar.
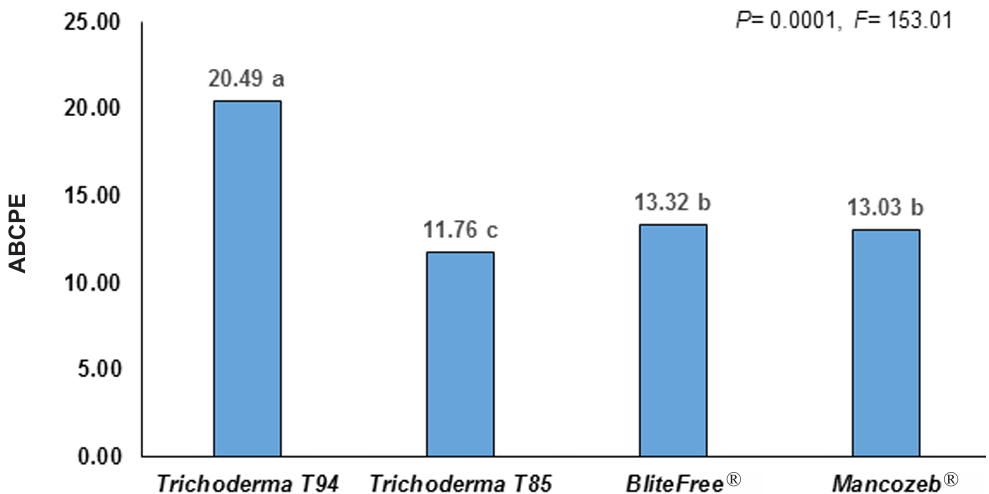
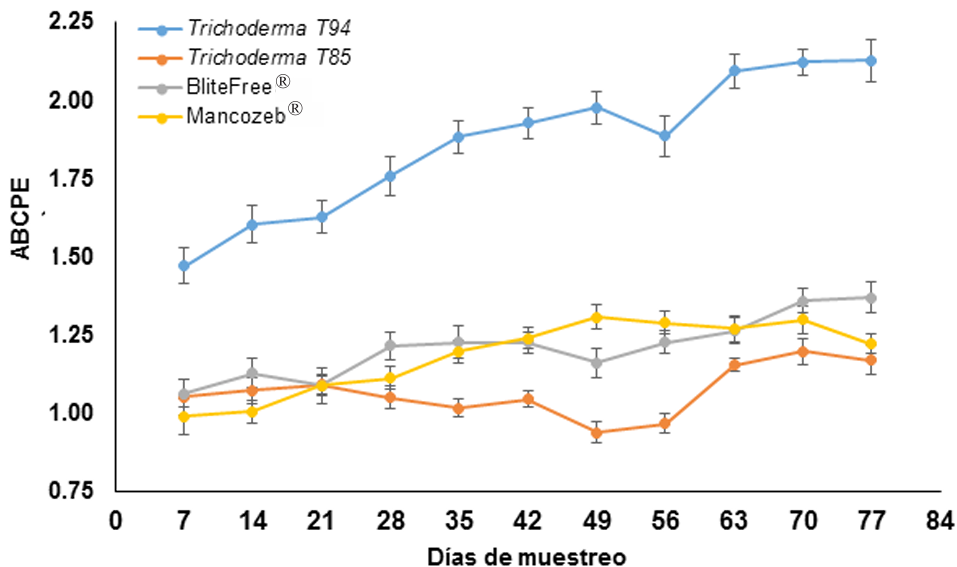
Resultados. El MQ incrementó el rendimiento de Trichoderma spp. en 71%, las cepas T-94 (1.41×108 conidiosporas g-1) y T-85 (1.20×108 conidiosporas g-1) presentaron los mayores rendimientos. La cepa T-85 redujo la severidad, PPI y ABCPE de la SN comparado con las aplicaciones del control químico “Mancozeb”.
Conclusión. El MQ fue el mejor sustrato para obtener mayor rendimiento en las cepas T-94 y T-85 de Trichoderma spp. La aplicación foliar semanal de conidiosporas de Trichoderma T-85 redujo la severidad, PPI y ABCPE de la SN en banano cv. Gran enano.
-
O copiar el link

PorJeny Michua Cedillo, Gustavo Mora Aguilera*, Gerardo Acevedo Sánchez
Recibido: 30/7/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2406-7
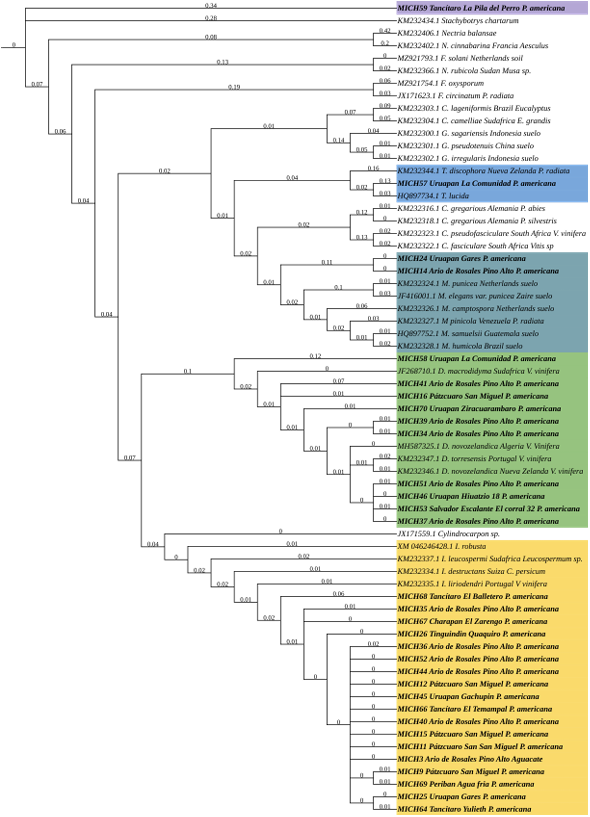
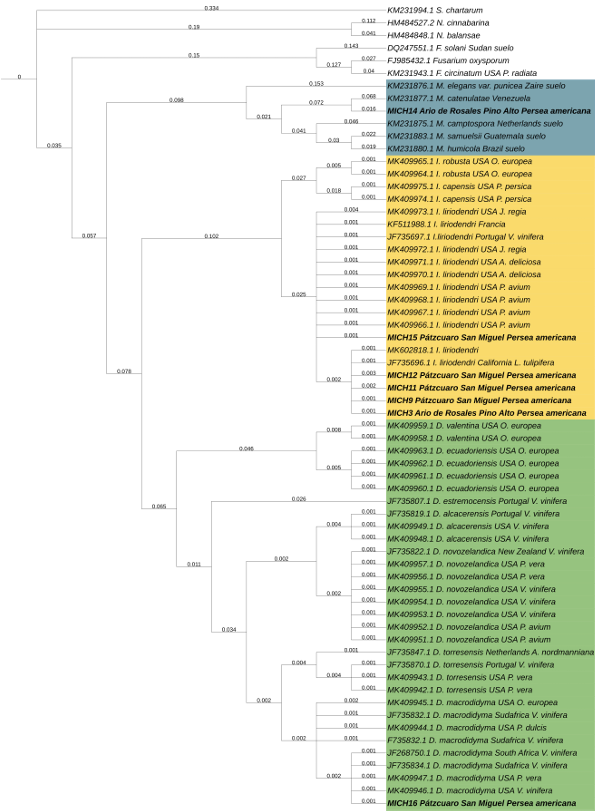
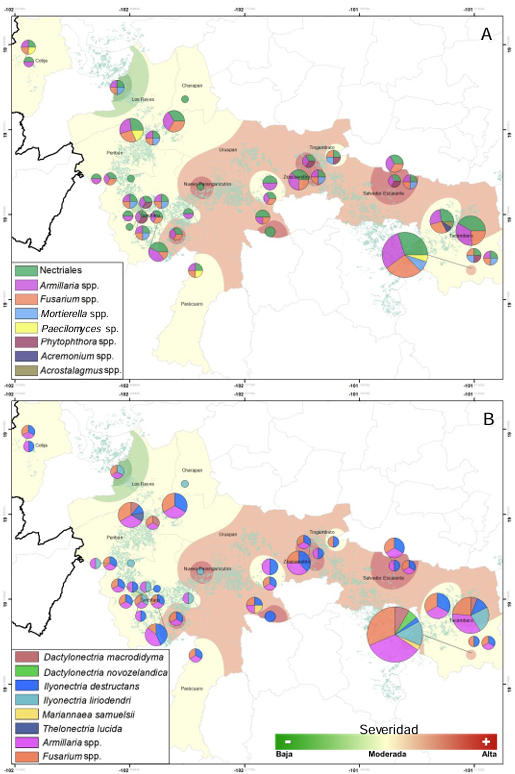
Resumen Antecedentes/Objetivo. Miembros de Nectriaceae se han detectado en Michoacán desde 2019. Sin embargo, su detección no ha tenido el enfoque etiológico primario por lo que se desconoce la identidad de especies en raíz, distribución geográfica y asociación con otras familias. El objetivo fue caracterizar biológica y molecularmente especies nectriales asociadas al aguacate (Persea americana).
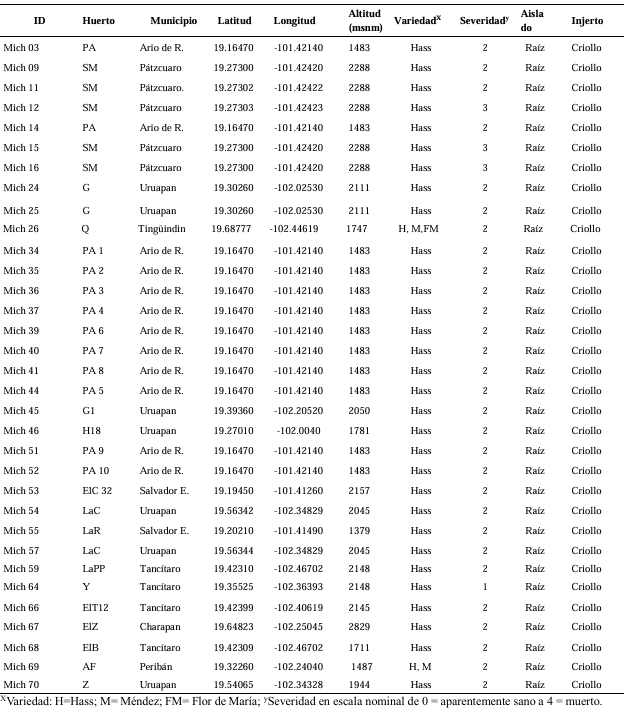
Materiales y Métodos. Se procesaron 70 muestras compuestas de raíces provenientes de árboles de aguacate con marchitez detectados en huertos comerciales de 13 municipios de Michoacán. Treinta aislados seleccionados con criterios epidemiológicos se cultivaron en extracto malta-agar, PDA y avena-agar para caracterización cultural y morfológica. A partir del ADN micelial se amplificaron los genes TEF 1-a y RPB2, las secuencias se limpiaron y alinearon con Seqassement y MAFFT, respectivamente. Se aplicaron algoritmos filogenéticos de inferencia Bayesiana y máxima parsimonia mediante PAUP4.0 y MrBayes3.2 complementándose con 66 y 65 secuencias del GenBank para TEF 1-a y RPB2, respectivamente. Cuatro especies de Hypocreales y S. chartarum se emplearon como especies externas.
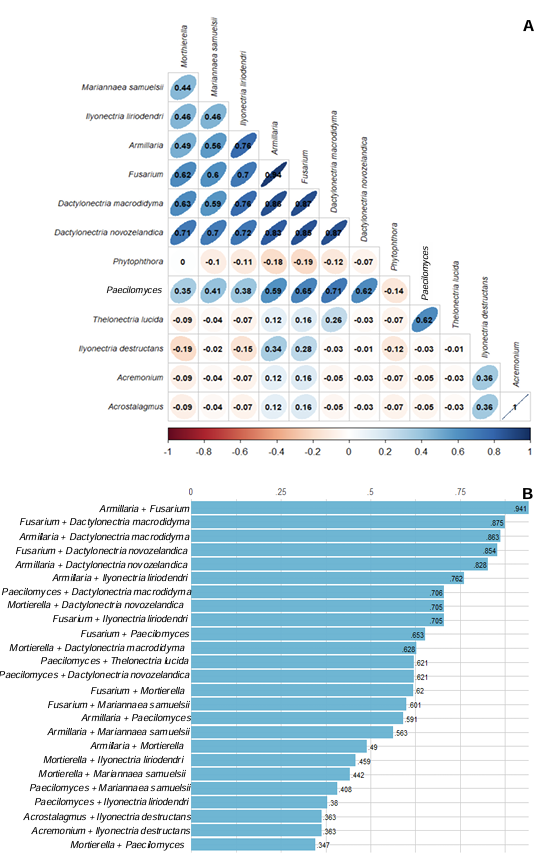
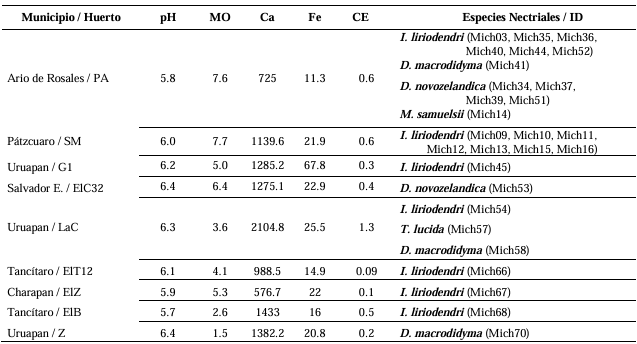
Resultados. Se obtuvieron cinco morfotipos de nectriales con crecimiento radial variable y coloración marrón. Se identificaron tres géneros y tres especies con TEF 1-a (>97% homología) y tres géneros y cinco especies con RPB2 (>97% homología) correspondientes a Ilyonectria (56% de los aislados), Dactylonectria (33 %), Mariannaea (6 %) y Thelonectria (3 %). En raíz, con niveles de daño variable, se observaron asociaciones significativas (p ≤ 0.05) entre especies nectriales con Armillaria (97.1 %), Fusarium (92.9 %), Paecilomyces (56.4 %) y Mortierella (47.3 %), no así con Phytophthora (r < - 0.07). Regionalmente, Ilyonectria liriodendry fue las más prevalente, preponderantemente asociada con Fusarium y/o Armillaria. El sur del municipio de Tacámbaro tuvo la mayor diversidad de especies nectriales y géneros fungosos en general.
Conclusión. Se postula la presencia regional de un síndrome de declinamiento en aguacate asociado a un complejo de hongos caracterizado por defoliación descendente, marchitez, reducción de crecimiento de fruto, decoloración y necrosis medular y cortical en raíz secundaria. Los nectriales Dactylonectria macrodidyma, D. novo-zelandica, Thelonectria lucida, Mariannaea samuelsii y I. liriodendry se asociaron con estos síntomas posiblemente con capacidad primaria de infección, pero frecuentemente en coinfección con Fusarium spp. y/o Armillaria spp. (r = 0.60 – 0.88, p ≤ 0.05). Otros hongos podrían estar asociados. Phytophthora podría tener limitada implicación en este síndrome. Este constituye el primer reporte de Ilyonectria, Dactylonectria, Mariannaea y Thelonectria asociados al aguacate (Persea americana) en México.
-
O copiar el link


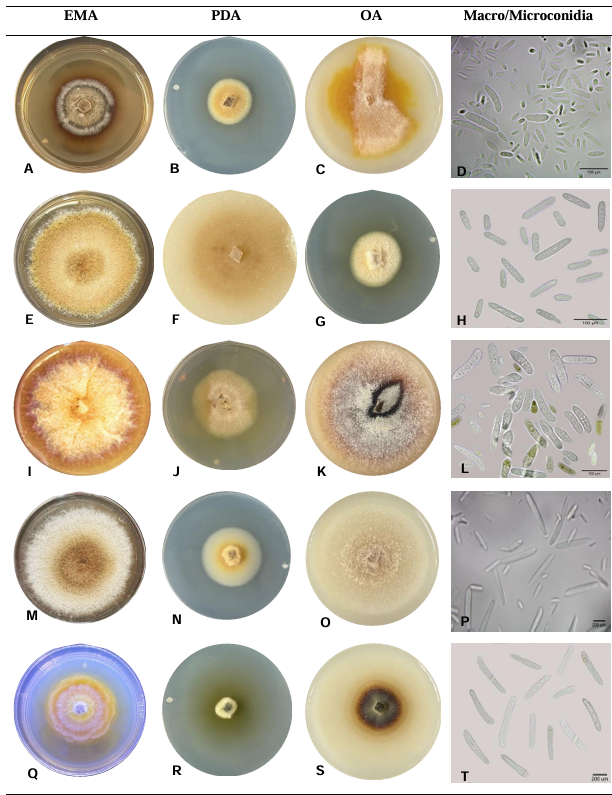
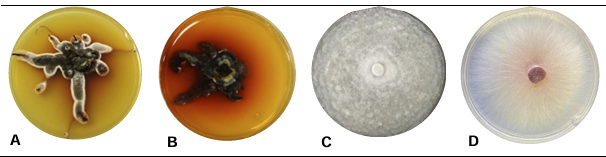
PorGabriela Olivares Rodriguez, Juan Gabriel Angeles Núñez, Francisco Mondragón Rojas, Patricia Rivas Valencia, José Luis Zárate Castrejón, Luis Antonio Mariscal Amaro, Luis Febronio Díaz Espino, Talina Olivia Martínez Martínez*
Recibido: 10/7/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-28
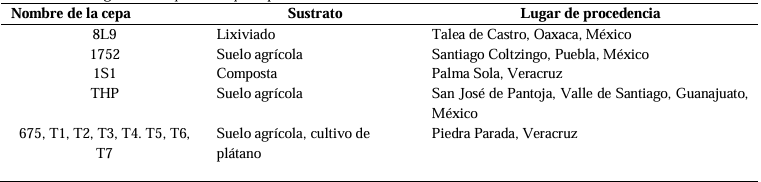
Resumen Antecedentes/Objetivo. El hongo Neopestalotiopsis sp. es un patógeno emergente que puede ocasionar pérdidas superiores al 70 % en el cultivo de fresa. Debido a esta situación, se busca implementar métodos de control de bajo impacto ecológico. El objetivo de este trabajo fue evaluar el efecto inhibitorio de cepas de Trichoderma sp. y de fungicidas empleados en el Bajío de Guanajuato, México, sobre el crecimiento de Neopestalotiopsis sp.
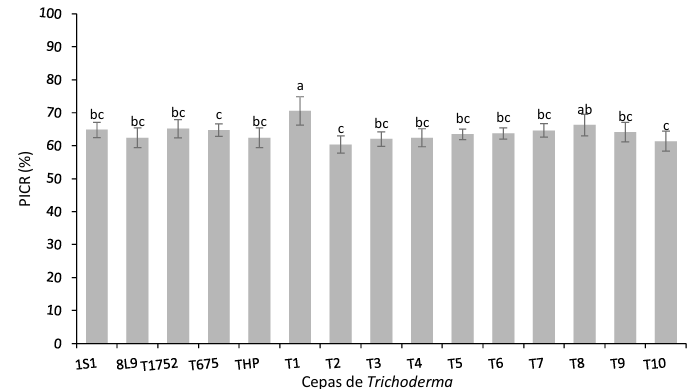
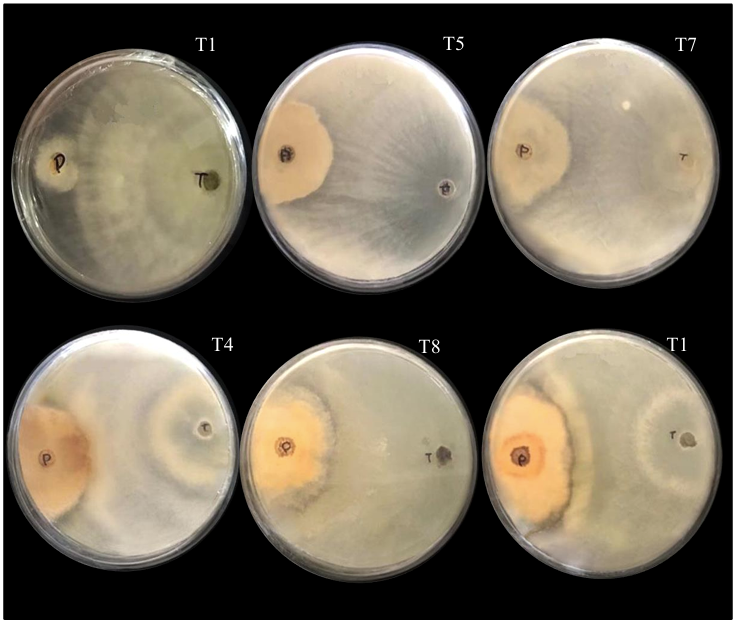
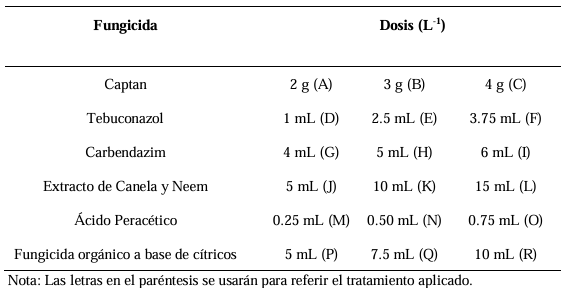
Materiales y Métodos. El patógeno se aisló de plantas de fresa sintomáticas. Se realizó la caracterización morfológica y de patogenicidad del aislado. Las cepas de Trichoderma se obtuvieron de la colección biológica del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias Campo Experimental Bajío (CEBAJ) y se confrontaron en cultivos duales con el patógeno, se determinó el porcentaje de inhibición del crecimiento radial (PICR) a las 120 h. Además, se evaluó el efecto de cinco fungicidas comerciales, que se agregaron al medio de crecimiento, sobre el diámetro de crecimiento del hongo.
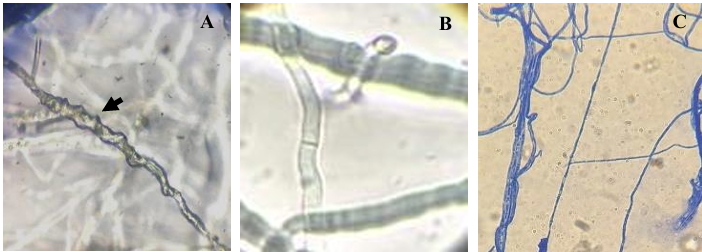
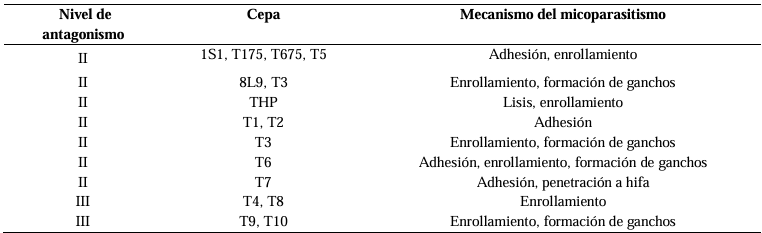
Resultados. Se obtuvo PICR en un intervalo de 63 al 70 %. El mecanismo de parasitismo de Trichoderma fue por enrollamiento, adhesión y lisis a las hifas del patógeno. La cepa T1 fue la que presentó mayor potencial para el control del patógeno, seguido de T5 y T7. Tres fungicidas comerciales, Tebuconazol (100 mL 100 L-1), Extracto de Canela y Neem (500 mL 100 L-1), y el Ácido Peracético (25 mL 100 L-1) inhibieron completamente el crecimiento del hongo.
Conclusión. Estos resultados contribuyen al conocimiento sobre el control de Neopestalotiopsis sp. con la aplicación de Trichoderma y los productos autorizados en México.
-
O copiar el link



PorAna Patricia Ibarra Valenzuela, Rosalba Troncoso Rojas, Alma Rosa Islas Rubio, Elizabeth Peralta, Herlinda Soto Valdez*, Hayati Samsudin
Recibido: 09/4/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-4
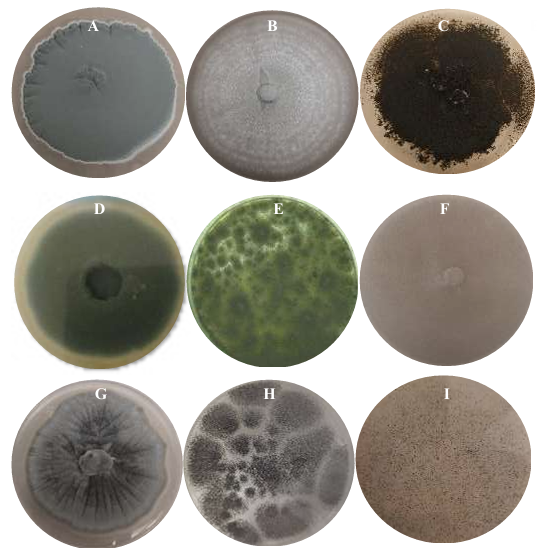
Resumen Antecedentes/Objetivo. Las tortillas de maíz, alimento básico en México, presentan una vida útil de 1-2 días a 25 °C debido al crecimiento fúngico. Una alternativa para extender la vida de anaquel de las tortillas es adicionar aceite esencial de clavo (AEC), sus componentes mayoritarios: eugenol (E), isoeugenol (I) y acetato de eugenilo (AE). El objetivo fue evaluar el efecto antifúngico del AEC sobre hongos identificados en tortillas de maíz.
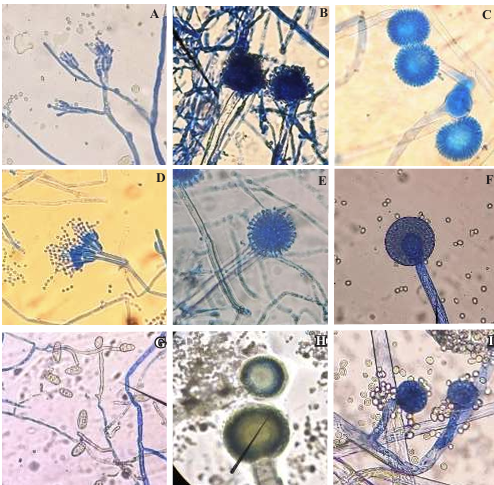
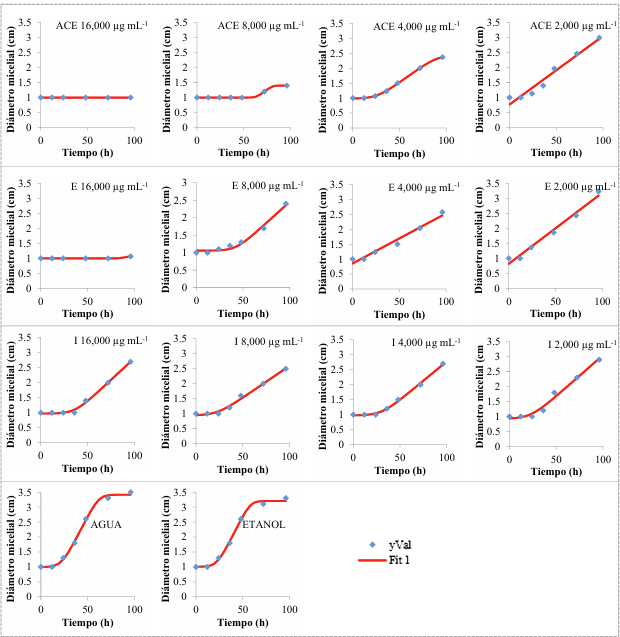
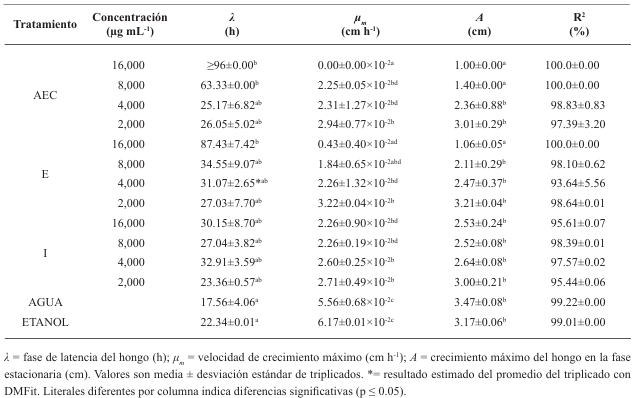
Materiales y Métodos. Se adquirieron muestras de un kg de tortillas de maíz de las capitales de cinco estados del país (Sonora, Nuevo León, Michoacán, Oaxaca y Yucatán). Los hongos se identificaron por su morfología y por biología molecular. Además, se les determinó la concentración mínima inhibitoria (CMI) frente a AEC. El efecto de E, I y AE sobre Aspergillus niger (previamente identificado) se evaluó con el modelo de Gompertz.
Resultados. Se obtuvieron dos aislados fúngicos de las tortillas de Nuevo León, Sonora, Yucatán y Michoacán y un aislado de Oaxaca, mismos que se identificaron por biología molecular: Aspergillus longivesica y Curvularia spicifera de Nuevo León; Aspergillus niger y Penicillium brevicompactum de Sonora; Aspergillus sp. de Oaxaca; Mucor sp. y Aspergillus flavus de Yucatán; Penicillium herquei, y Curvularia racemosus de Michoacán. Las CMIs fueron 200, 400, 800, 400, 800, 400, 800, 800 y 400 µg mL-1, respectivamente. AEC, E e I a 800 µg mL-1 retardaron la fase exponencial de crecimiento de Aspergillus niger, mientras que AE no mostró efecto.
Conclusión. El AEC podría ser una alternativa natural para prolongar la vida útil de tortillas de maíz.
-
O copiar el link


PorAugusto Gil Ceballos Ceballos, Yisa María Ochoa Fuentes*, Ernesto Cerna Chávez, Arely Cano García
Recibido: 18/4/2024 – Publicado: 31/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-5
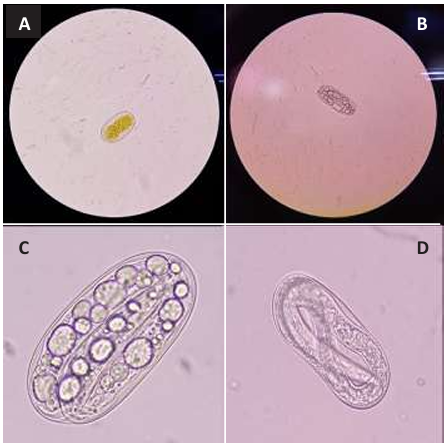
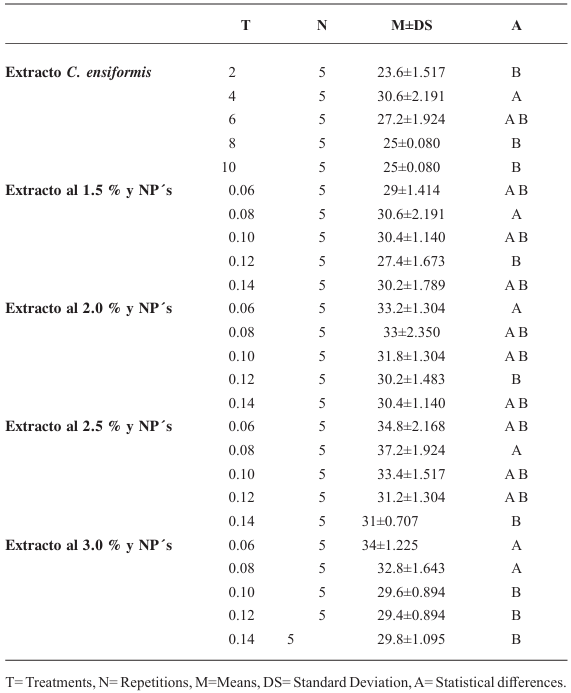
Resumen Antecedentes/Objetivo. Los extractos de semillas de Canavalia ensiformis han demostrado efectos antiparasitarios y repelentes contra plagas. El objetivo fue evaluar la efectividad del extracto combinado con nanopartículas (NP's) de dióxido de silicio contra los huevos de Meloidogyne incognita.
Materiales y Métodos. Se llevaron a cabo experimentos in vitro para evaluar los efectos de los extractos de semillas de C. ensiformis, tanto por sí solos como combinados con NP's de dióxido de silicio, en la eclosión de juveniles de M. incognita. Se utilizaron 150 huevos y se aplicaron concentraciones de 0, 2, 4, 6, 8 y 10 % del extracto. Además, se evaluaron concentraciones del extracto al 0, 1.5, 2.0, 2.5 y 3.0 %, cada una combinada con concentraciones de NP's al 0.06, 0.08, 0.10, 0.12 y 0.14 %.
Resultados. Ninguno de los tratamientos logró evitar la eclosión más del 30 % de juveniles. La modificación de la técnica de obtención del extracto de semillas de C. ensiformis puede causar un efecto ovicida complementario; sin embargo, al aumentar las concentraciones del extracto, puede propiciar la proliferación de hongos saprofitos y otros microorganismos.
Conclusión. Los tratamientos no mostraron efectos ovicidas significativos arriba del 40 %.
-
O copiar el link
Estimación de pérdidas provocadas por Potato virus Y en el cultivo de papa en Coahuila
PorJoel De Santiago Meza*, Gustavo Alberto Frías Treviño, Luis Alberto Aguirre Uribe, Alberto Flores Olivas
Recibido: 05/4/2024 – Publicado: 30/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-2
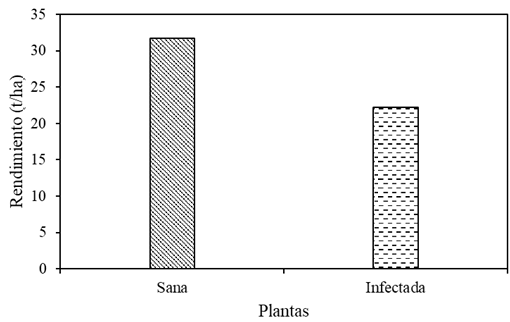
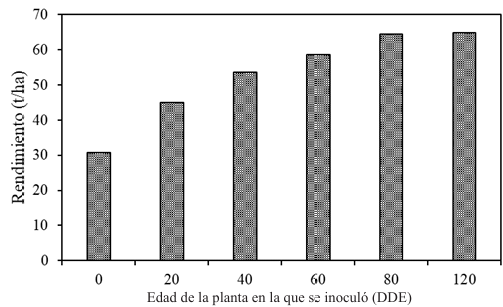
Resumen Antecedentes/Objetivo. El objetivo fue evaluar experimentalmente las pérdidas ocasionadas por PVY (Potato Virus Y) en el cultivo de papa variedad Fianna y, por consiguiente, estimar las pérdidas ocasionadas por este virus en la zona productora de papa de Coahuila.
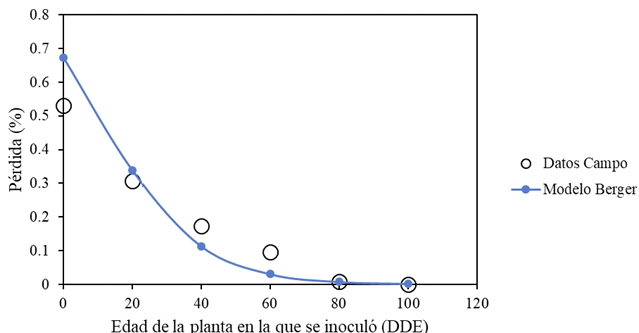
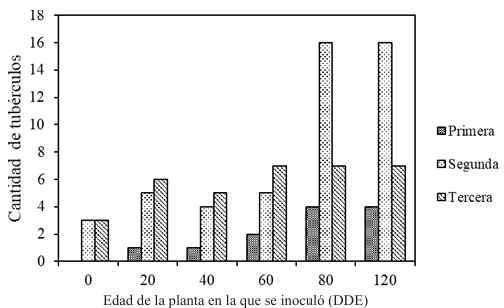
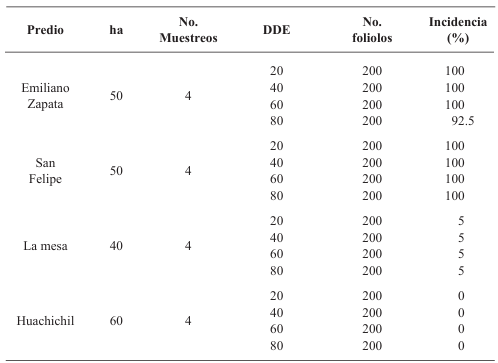
Materiales y Métodos. Surcos de una parcela experimental sembrados con plántula y semilla-tubérculo de papa, se inocularon mecánicamente con PVY a los 20, 40, 60 y 80 días después de la emergencia. Se cosecharon los tubérculos producidos y se evaluaron las pérdidas en cada tratamiento. Adicionalmente, en cuatro predios comerciales de papa en este mismo estado, se tomaron muestras de foliolos a los 20, 40, 60 y 80 días después de la emergencia, y se evaluó mediante pruebas de ELISA el porcentaje de plantas infectadas con PVY. Los datos de pérdidas de la parcela experimental y los datos de incidencia de los predios se utilizaron para elaborar un modelo estadístico para estimar las pérdidas causadas por PVY en la región de Coahuila.
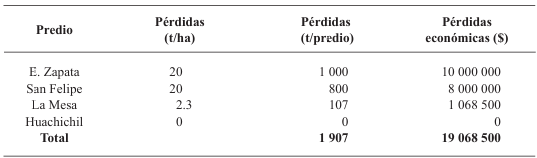
Resultados. Las pérdidas en el rendimiento por PVY en la parcela experimental fueron de 9.4% a 53%. El porcentaje de incidencia de plantas infectadas en los predios comerciales varió de 0% a 100%. El modelo que mejor se ajustó a los datos obtenidos fue el de Berger . Las pérdidas estimadas en la zona productora de papa de Coahuila en el ciclo 2022 fueron de 18 %, equivalente a $19, 068, 500.
Conclusión. Esta información resalta la importancia de utilizar semilla certificada libre de PVY y de proteger el cultivo desde la emergencia hasta los 60 DDE.
-
O copiar el link


PorAlejandra Chacón López, José Luis Hernández Flores, Efigenia Montalvo González, Selene Aguilera Aguirre*
Recibido: 20/8/2024 – Publicado: 30/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2308-2
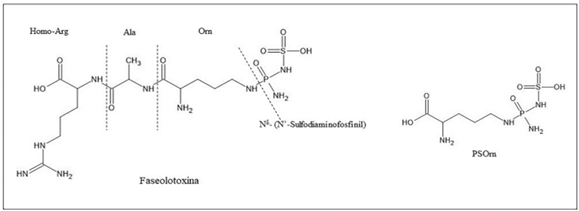
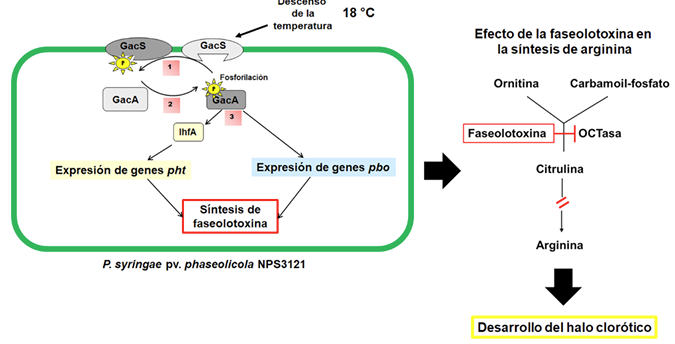
Resumen Antecedentes/Objetivo. La faseolotoxina es producida por uno de los fitopatógenos más importantes y estudiados en el área agrícola: Pseudomonas syringae pv. phaseolicola. Esta bacteria causa el tizón de halo, una enfermedad que devasta al cultivo del frijol. El éxito de P. syringae pv. phaseolicola está relacionado con su información genética que le permite sintetizar metabolitos deletéreos para su hospedero, como la faseolotoxina. El objetivo de la presente investigación fue analizar la base molecular del mecanismo de acción, la inmunidad, la genética involucrada en la biosíntesis de la faseolotoxina, las estrategias de diagnóstico molecular y las técnicas moleculares desarrolladas en México, para llevar a cabo el manejo del tizón de halo del frijol.
Materiales y Métodos. Se realizó la búsqueda y el análisis de la información científica más relevante respecto a la biosíntesis de faseolotoxina y los estudios moleculares de los factores de patogenicidad y virulencia de P. syringae pv. phaseolicola que han contribuido al desarrollo de estrategias moleculares enfocadas en el diagnóstico y manejo del tizón de halo en frijol.
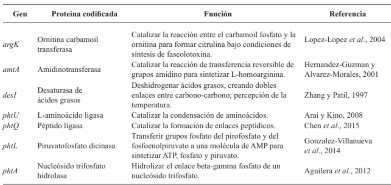
Resultados. P. syringae pv. phaseolicola produce faseolotoxina, que es la responsable de la formación del halo clorótico característico del tizón de halo, esta toxina es un inhibidor de la OCTasa, una enzima que participa en la ruta de síntesis de arginina en frijol. Las regiones cromosómicas Pht y Pbo contienen genes involucrados en la síntesis e inmunidad de la faseolotoxina, y la expresión de estos genes está regulada por el sistema GacS/GacA y la temperatura. La identificación de genes involucrados en la síntesis de factores de patogenicidad y virulencia, como la faseolotoxina, ha permitido el desarrollo de estrategias de diagnóstico y manejo de la enfermedad basadas en la amplificación de ADN y el uso de marcadores moleculares que facilitan la identificación de cultivares de frijol resistentes al patógeno.
Conclusión. Los estudios moleculares han contribuido al entendimiento de cómo el patovar phaseolicola produce faseolotoxina. Esta información ha sido esencial para entender cómo las bacterias han evolucionado de variantes no patogénicas a patogénicas; además, proporcionan información que permite desarrollar nuevas estrategias para un diagnóstico oportuno y contribuyen en las estrategias para el manejo del tizón de halo.
-
O copiar el link




PorConrado Parraguirre Lezama, Omar Romero Arenas*, Alba Cruz Coronel, Amparo Mauricio Gutiérrez, Carlos A Contreras Pare, Antonio Rivera Tapia
Recibido: 25/2/2024 – Publicado: 27/12/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2402-7
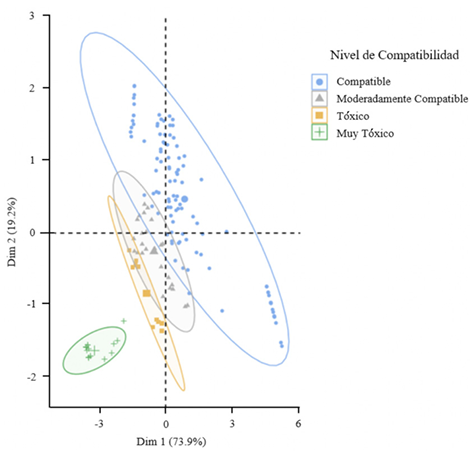
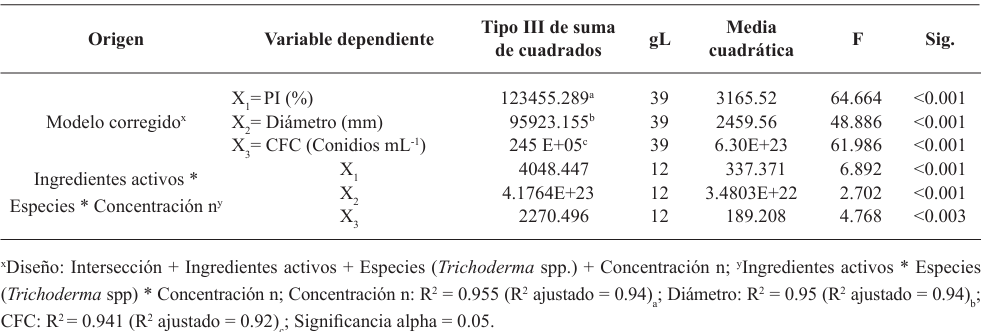
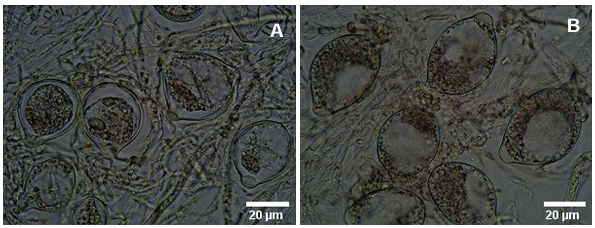
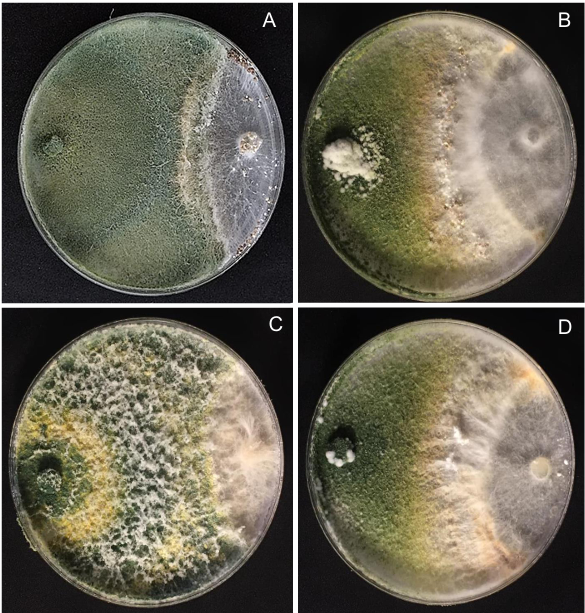
Resumen Objetivo/Antecedentes. La transición hacia prácticas agrícolas responsables es fundamental para promover la salud de los agroecosistemas y garantizar la seguridad alimentaria. Impulsar investigaciones integrales que combinen métodos químicos y biológicos representa un avance significativo en el manejo de fitopatógenos, es decir, esta aproximación novedosa se basa en la premisa de que la acción conjunta entre fungicidas y un agente antagónico como Trichoderma spp., pueden ofrecer una protección robusta en comparación con enfoques individuales. El objetivo del estudio es investigar la resistencia y compatibilidad in vitro de cuatro especies de Trichoderma frente a tres fungicidas ampliamente utilizados en México.
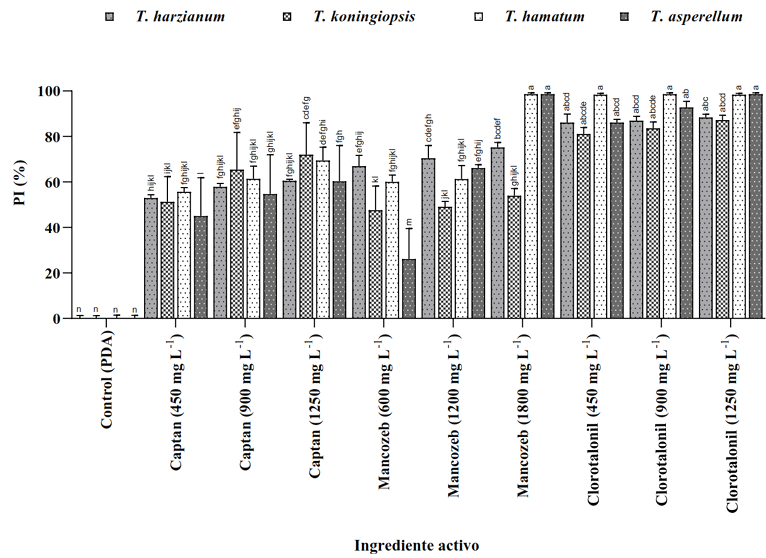
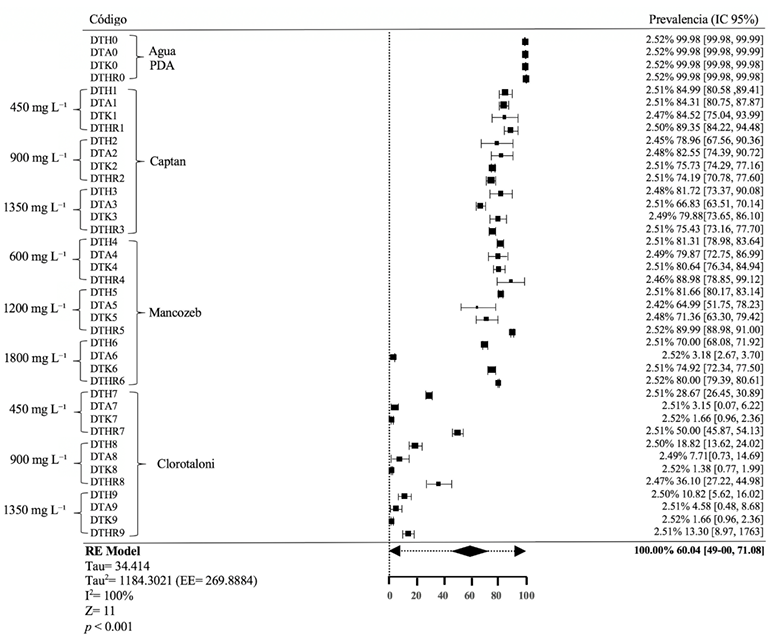
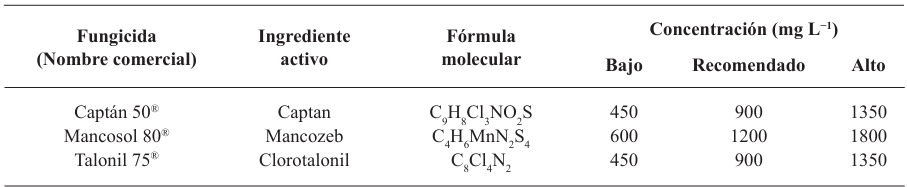
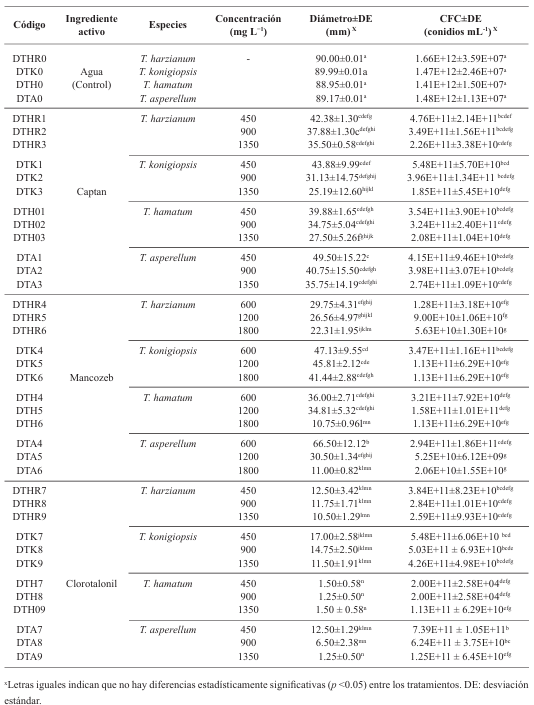
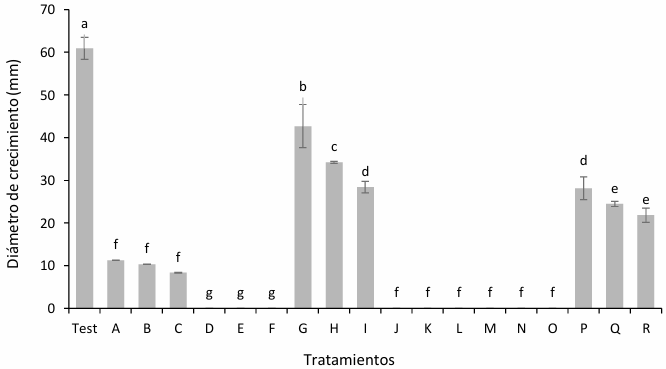
Materiales y Métodos. Se empleó la técnica de intoxicación controlada en medio PDA bajo condiciones controladas con tres concentraciones (450, 900 y 1350 mg L−1), pare el caso de los ingredientes activos Captan y Clorotalonil, para Mancozeb se utilizaron 600, 1200 y 1800 mg L−1. La compatibilidad se determinó en relación con el grupo control utilizando el software estadístico SPSS Statistics versión 26 para el entorno operativo Windows.
Resultados. El estudio reveló que las cepas de T. harzianum, T. hamatum, T. koningiopsis y T. asperellum exhibieron una compatibilidad global del 60.04% para los ingredientes activos evaluados, siendo el fungicida Captán 50® el que demostró el mayor porcentaje de compatibilidad (79.87%) en las concentraciones de 450, 900 y 1350 mg L–1. T. harzianum mostró mayor tolerancia al ingrediente activo Clorotalonil en la concentración de 450 mg L⁻¹, sin embargo, a concentraciones más altas demostró mayor toxicidad, siendo T. koningiopsis la que exhibió la menor resistencia en sus tres concentraciones evaluadas.
Conclusión. Los tratamientos con diferentes concentraciones de los fungicidas Captan, Mancozeb y Clorotalonil evidenciaron una marcada variabilidad en términos de prevalencia y toxicidad hacia las especies evaluadas de Trichoderma in vitro. Este enfoque permite diseñar estrategias de manejo integrado minimizando la dependencia de productos químicos y promoviendo la compatibilidad entre agentes biológicos y fungicidas.
-
O copiar el link