

-
O copiar el link


Filtros de Contribuciones
Buscar Contribución:
PorVíctor Manuel Rodríguez Romero, Ramón Villanueva Arce, Enrique Durán Páramo*
Recibido: 05/3/2024 – Publicado: 20/6/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2405-3
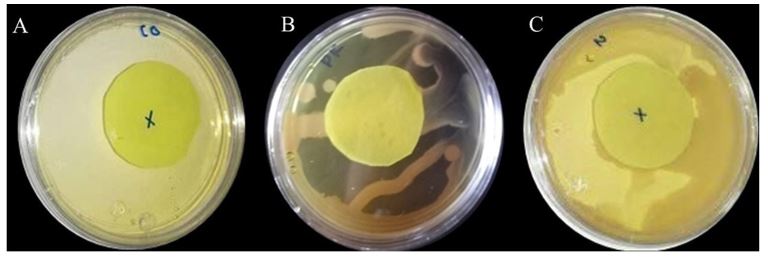
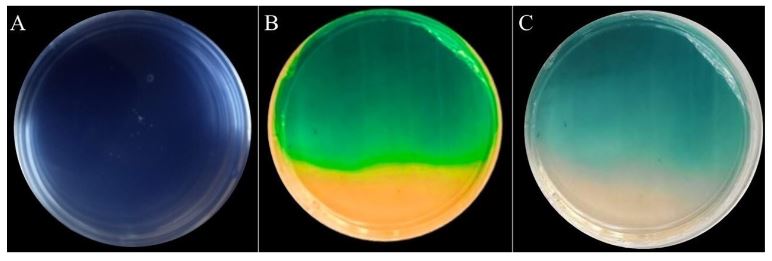
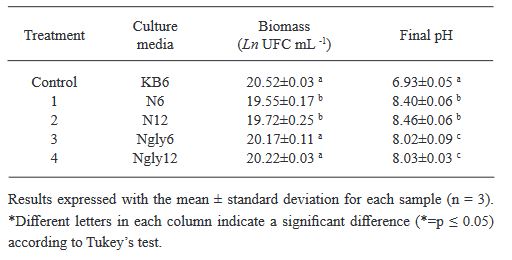
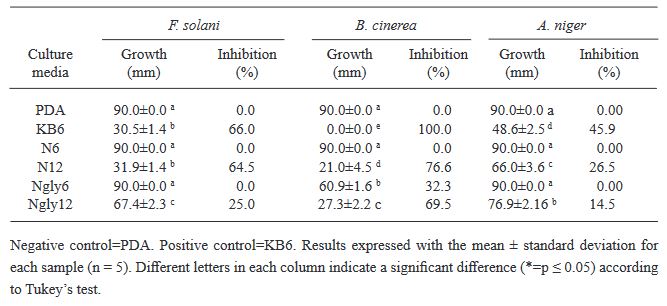
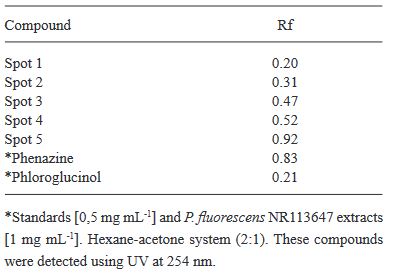
Resumen Antecedentes/Objetivo. El nejayote es un residuo agroindustrial alcalino que se genera a partir del proceso de nixtamalización del maíz. El propósito de este trabajo fue demostrar que el nejayote puede ser utilizado como medio de cultivo para el crecimiento de Pseudomonas fluorescens NR113647 y para la producción de metabolitos con actividad antifúngica para el manejo sustentable de Aspergillus niger, Botrytis cinerea y Fusarium solani.
Materiales y Métodos. Se formularon medios de cultivo con nejayote y nejayote con glicerol, con pH 6 y 12. La biomasa bacteriana fue separada por centrifugación y filtración y se determinó la capacidad antifúngica in vitro de los extractos contra A. niger, B. cinerea y F. solani. Además, se realizó la determinación de los metabolitos presentes en los extractos. P. fluorescens NR113647 fue capaz de crecer en todos los medios.
Resultados. Los extractos provenientes de nejayote a pH 12 mostraron inhibición del crecimiento de todos los hongos evaluados; se identificaron al menos cinco metabolitos producidos por P. fluorescens NR113647 involucrados en el biocontrol de fitopatógenos.
Conclusión. El nejayote puede ser usado como medio de cultivo para P. fluorescens NR113647, para la producción de biomasa y metabolitos secundarios con capacidad antifúngica; además, el nejayote podría usarse para el cultivo de otros microorganismos.
-
O copiar el link
PorWilliam Villalobos Muller, Laura Garita Salazar, Ana María Conejo Salazar, Izayana Sandoval Carvajal, Mauricio Montero Astúa, Lisela Moreira Carmona*
Recibido: 05/3/2024 – Publicado: 20/6/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2403-1
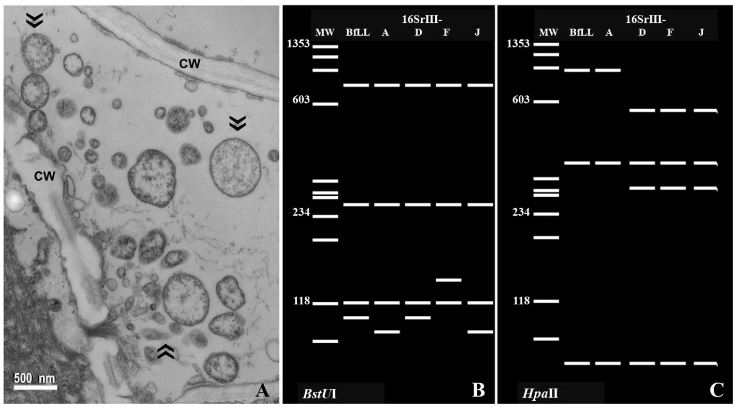
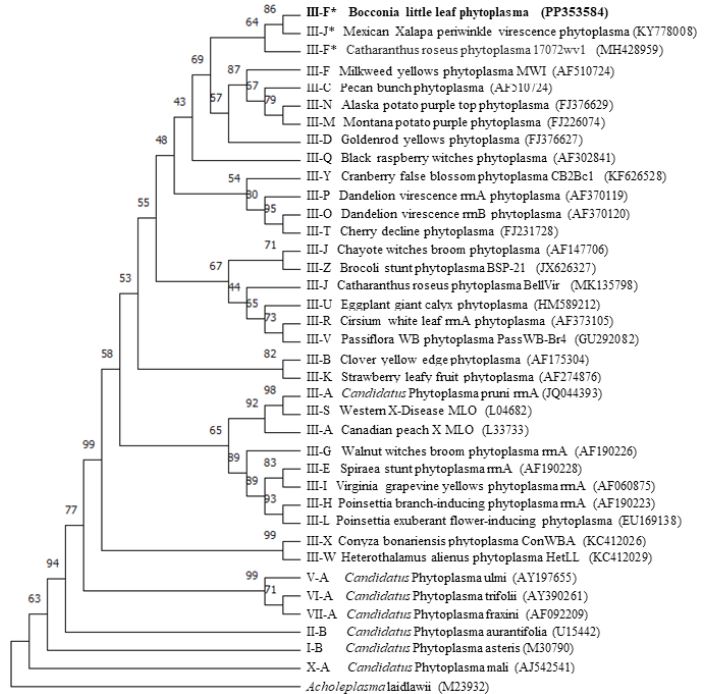
Resumen Antecedentes/Objetivo. Bocconia frutescens (Papaveraceae) es un árbol pequeño, distribuido naturalmente desde México hasta Argentina y en la cuenca del Caribe. Árboles de Bocconia con síntomas semejantes a los inducidos por fitoplasmas, como hoja pequeña y escobas de bruja, fueron encontrados en la provincia de Cartago, Costa Rica. Detectar e identificar al posible fitoplasma asociado con dichos síntomas fue el objetivo de este estudio.
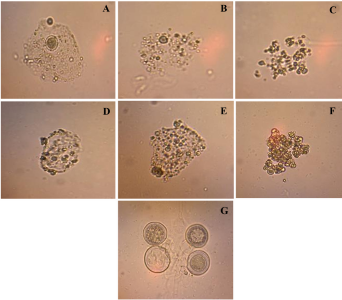
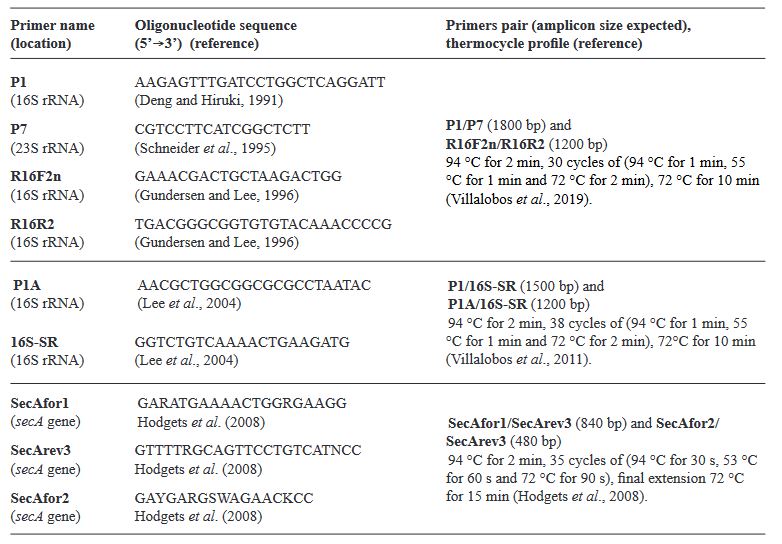
Materiales y Métodos. Se evaluó tejido foliar empleando microscopia electrónica de transmisión (TEM), PCR anidada empleando cebadores universales y específicos para amplificar los genes ARNr 16S, y secA de fitoplasmas. Las secuencias nucleotídicas (método Sanger) obtenidas de los amplicones, se emplearon para BLASTn, análisis filogenéticos y RFLP’s in silico.
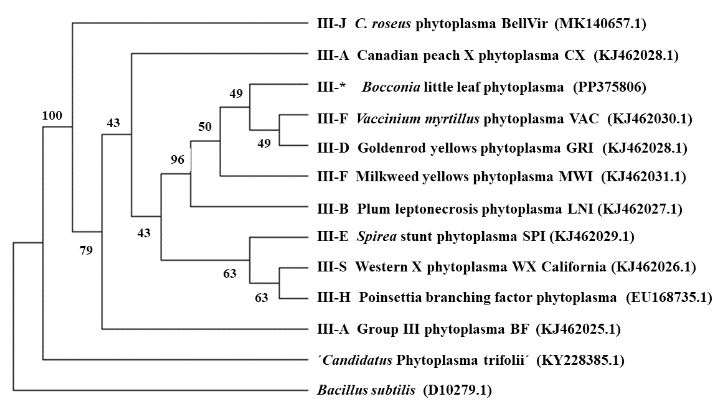
Resultados. La presencia de fitoplasmas en el tejido del floema se observó mediante TEM solamente en los árboles sintomáticos. Los diferentes análisis con las secuencias parciales (genes 16Sr y secA) indicaron la presencia de una cepa relacionada al Candidatus Phytoplasma pruni en las muestras evaluadas.
Conclusión. Fitoplasmas se encontraron sólo en los árboles sintomáticos de Bocconia evaluados. Los fitoplasmas se identificaron como una cepa relacionada al ´Ca. Phytoplasma pruni´. Este es el primer reporte de B. frutescens como un hospedero natural de Ca . Phytoplasma pruni.
-
O copiar el link

Identificación de fitoplasmas asociados al Bunchy Top de la papaya en Colima, México
PorPedro Valadez Ramírez, Daniel Leobardo Ochoa Martínez*, Guadalupe Valdovinos Ponce, Edith Blanco Rodríguez, Sergio Aranda Ocampo, Candelario Ortega Acosta, Marco Tulio Buenrostro Nava, Jetzajary Ayerim Rodríguez Barajas, Luis Rafael De la Torre Velázquez, Carlos Luis Leopardi Verde
Recibido: 05/3/2024 – Publicado: 18/6/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2403-2
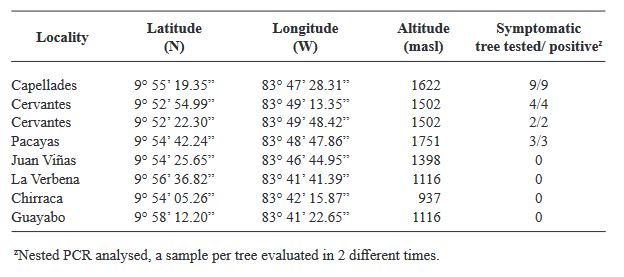
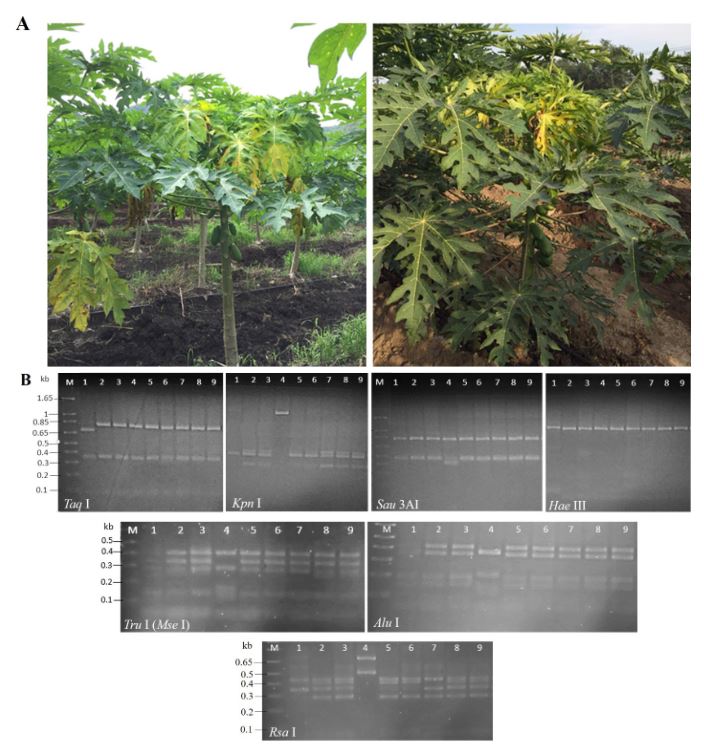
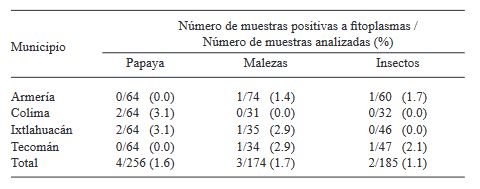
Resumen Antecedentes/Objetivo. Fitoplasmas, rickettsias y virus se han detectado en plantas de papaya con la enfermedad Bunchy Top (BT). En agroecosistemas de papaya ubicados en Colima, México, durante 2019 se observaron plantas con síntomas similares al BT. Con la finalidad de determinar la presencia de fitoplasmas e identificar las especies y/o subgrupos asociados a esta enfermedad, se recolectaron plantas asintomáticas y con síntomas en cuatro municipios productores del estado, así como malezas e insectos asociados al cultivo.
Materiales y Métodos. La detección e identificación de fitoplasmas se hizo a través de PCR, secuenciación y análisis filogenéticos de la subunidad SecA de la translocasa (secA) y 16S RNA ribosomal (16Sr), y PCR-RFLPs in vitro e in silico del gen 16Sr.
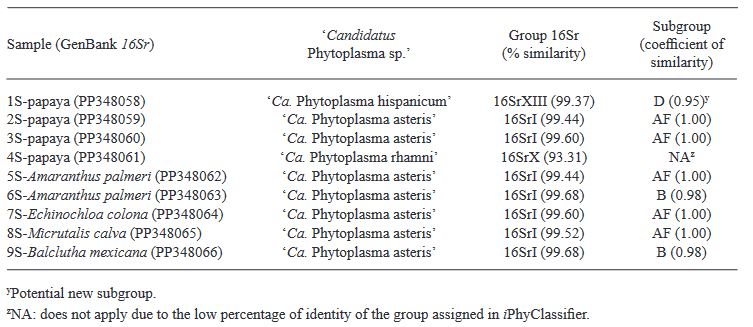
Resultados. En papaya se identificaron fitoplasmas de los grupos 16SrI (subgrupo AF), 16SrX y 16SrXIII, en el 2.08% (4 de 192) de las muestras con síntomas. Los resultados del análisis RFLPs in silico del gen 16Sr mostraron la presencia de los (sub)grupos 16SrX y 16SrXIII. En malezas e insectos, se identificaron fitoplasmas del grupo 16SrI (subgrupos AF y B) en el 1.7% (3 de 174) y 1.1% (2 de 185) de las muestras analizadas, respectivamente. Las malezas portadoras de fitoplasmas fueron Amaranthus palmeri y Echinochloa colona, mientras que los insectos fueron Micrutalis calva y Balclutha mexicana. Se reporta por primera vez la presencia de fitoplasmas 16SrI-AF, 16SrX y 16SrXIII en plantas de papaya con síntomas de Bunchy Top en agroecosistemas del estado de Colima, México. Los fitoplasmas de los grupos 16SrX y 16SrXIII se registran por primera vez a nivel mundial y en México en papaya, respectivamente. Las malezas e insectos portadores de fitoplasmas constituyen nuevos registros como reservorios naturales y potenciales vectores de las bacterias.
-
O copiar el link


PorMaría Emilia Belingheri Lagunes, Rosario Medel Ortiz*, Alejandro Salinas Castro, Dora Trejo Aguilar
Recibido: 15/1/2024 – Publicado: 07/6/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2401-1
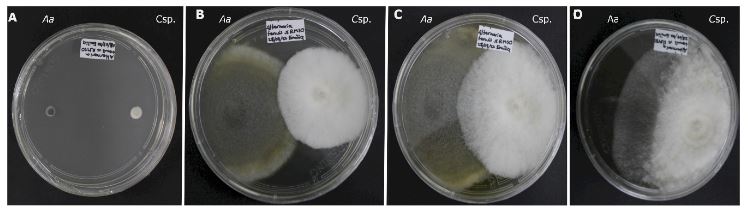
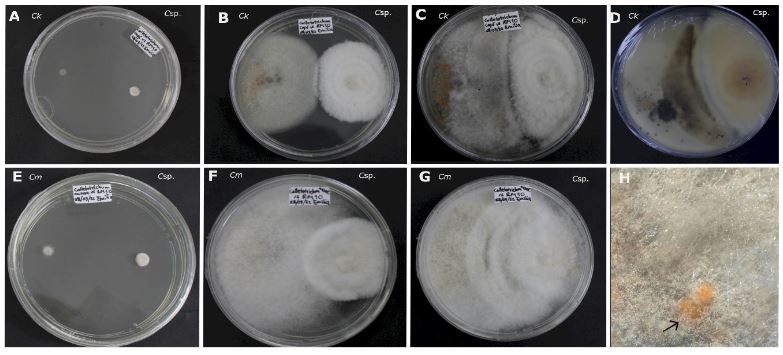
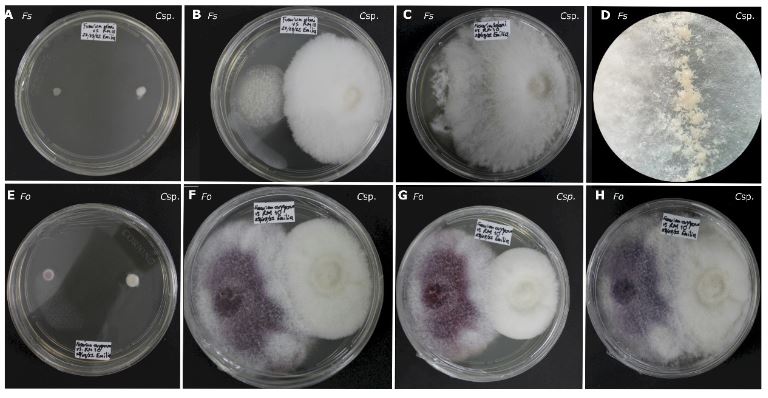
Resumen Antecedentes/Objetivo. El objetivo de este trabajo fue evaluar la capacidad antagónica in vitro de una cepa de Clonostachys sp., contra cinco especies de hongos asociados a enfermedades en cultivos de importancia económica.
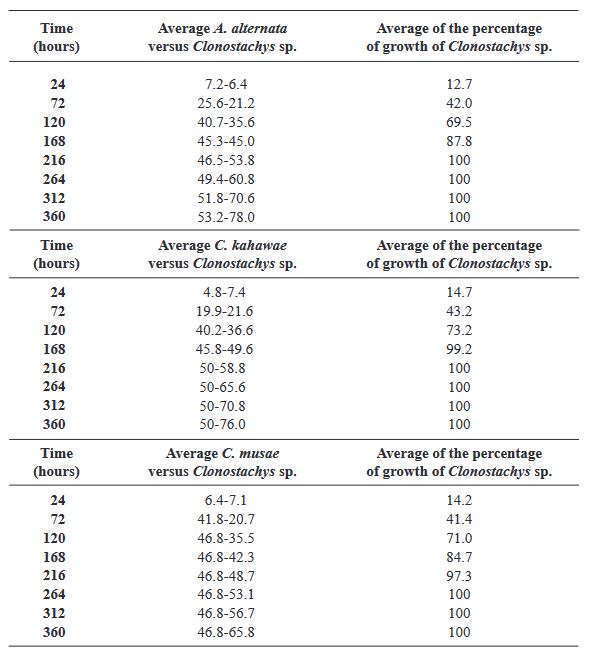
Materiales y Métodos. Se probaron cinco especies de hongos asociados a enfermedades de cultivos: Alternaria alternata, Colletotrichum kahawae, C. musae, Fusarium oxysporum y F. solani. Se realizaron cultivos duales con cinco repeticiones mas los controles. Se registró el crecimiento cada 24 horas, hasta completar 360 horas. Se determinaron las interacciones, el grado de antagonismo y se calculó el porcentaje de colonización. Los análisis estadísticos se realizaron con un modelo lineal generalizado (GLM).
Resultados. Todas las especies evaluadas mostaron antagonsimo del tipo sobrecrecimiento. El grado de antagonismo se clasificó en tres clases, siendo la clase dos la que se presentó en tres de las especies. El porcentaje de colonización fue del 100 % a las 216 h en tres de las especies y de 264 h para las otras dos. No hubo diferencias significativas en el porcentaje de colonización (p =0.0073), pero sí en el tiempo de invasión (p< 0.0001).
Conclusión. Los ensayos duales para comprobar el efecto antagónico in vitro constituyen la base para la selección de candidatos para el control biológico de hongos.
-
O copiar el link

PorAlejo Jairo Cristóbal*, José María Tun Suárez, Arturo Reyes Ramírez, Alberto Uc Várguez, Silvia Edith García Díaz
Recibido: 01/5/2024 – Publicado: 12/7/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2405-5
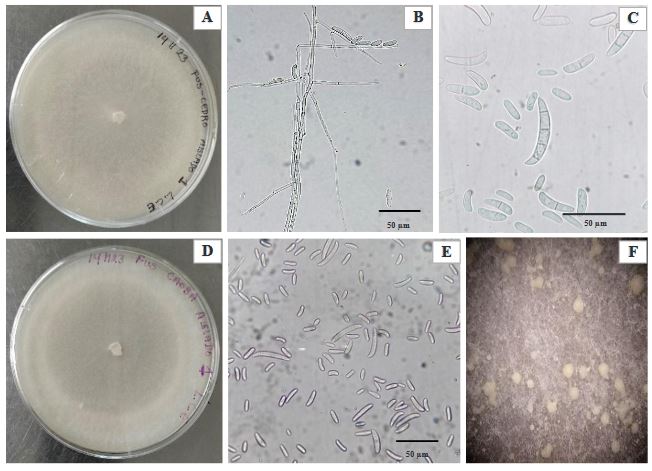
Resumen Antecedentes/Objetivo. En el estado de Yucatán, México, en los últimos cinco años se produjeron 10 millones de plantas forestales para diversas acciones de conservación y restauración. Las principales limitantes en la producción de estas plantas en vivero, son las enfermedades inducidas por Fusarium spp., que causan pudriciones de tallo y raíz y pérdidas de producción de plantas hasta el 50%. El objetivo del trabajo fue identificar el agente causal asociado con la pudrición y necrosis de tallo y raíz de cedro (Cedrela odorata) y caoba (Swietenia macrophylla) y su sensibilidad in vitro a fungicidas convencionales.
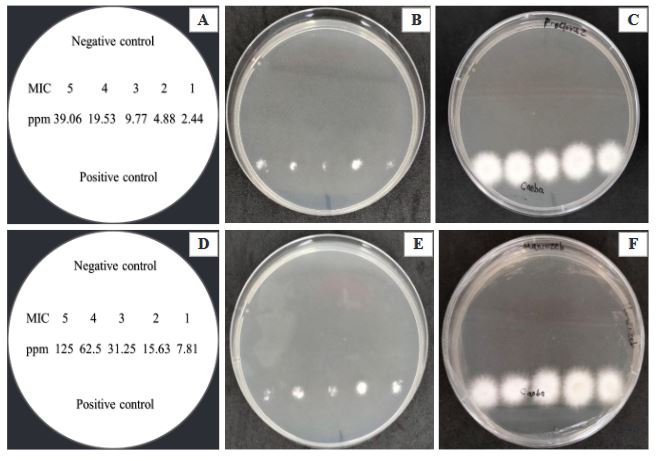
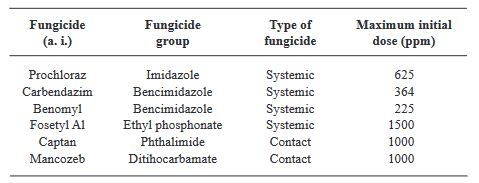
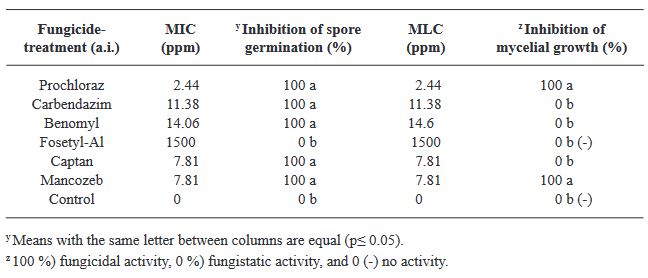
Materiales y Métodos. Se colectaron plantas de C. odorata y S. macrophylla de tres y seis semanas de germinación, respectivamente, con síntomas de necrosis y pudrición indicados; de donde se obtuvieron cinco aislados fúngicos y se identificaron morfológica y molecularmente. Se determinó in vitro por el método de microdilución, la Concentración Mínima Inhibitoria (CMI) de esporas y la Concentración Mínima Letal (CML) de seis fungicidas convencionales de aplicación recurrente en la región (Procloraz, Carbendazim, Benomilo, Fosetyl Al, Captan y Mancozeb) y validar su efectividad y viabilidad en el manejo de esta problemática.
Resultados. La morfología y secuencias moleculares de los aislados, fueron similares a las reportadas para Fusarium solani. La CMI de las esporas de F. solani para Procloraz, Carbendazim, Benomilo, Captan y Mancozeb fueron de 2.44. 11.38, 14.06, 7.81 y 7.81 ppm, respectivamente; Fosetyl Al, no inhibió la germinación de esporas ya que se observó crecimiento micelial normal del hongo en las concentraciones evaluadas.
Conclusión. El Procloraz y el Mancozeb, tuvieron la menor CML con 2.44 y 7.81 ppm, respectivamente.
-
O copiar el link

PorMiguel Ángel Ruíz González, Miguel Ángel Serrato Cruz*, Ernestina Valadez Moctezuma, Roney Solano Vidal
Recibido: 28/1/2024 – Publicado: 09/7/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2401-5
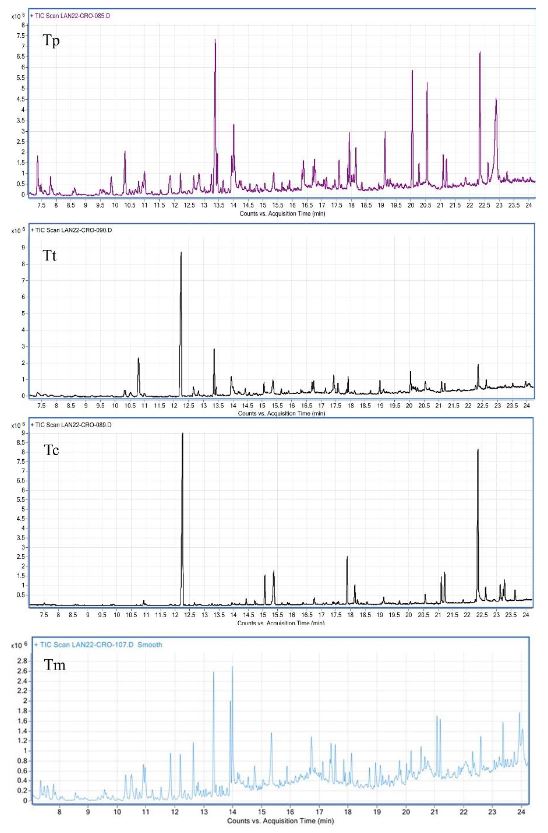
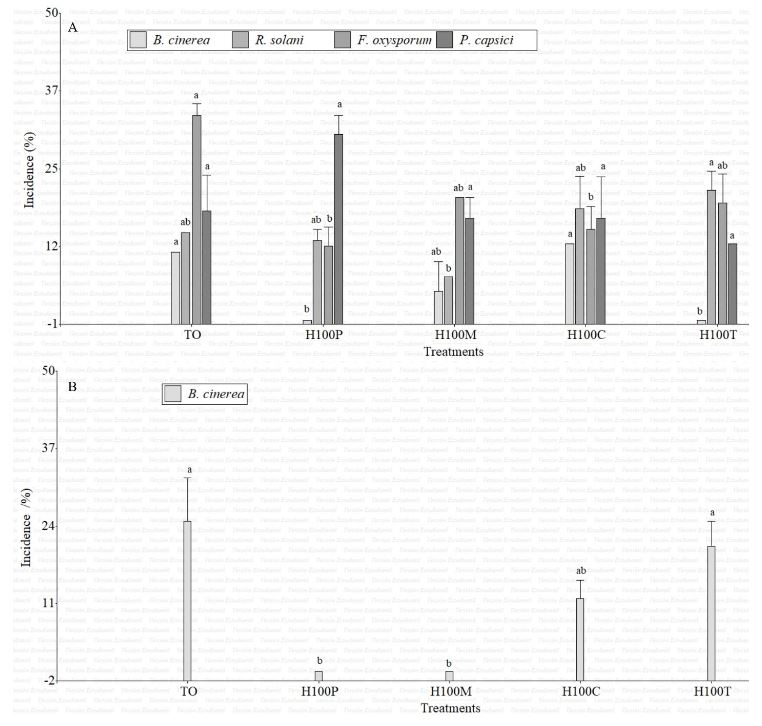
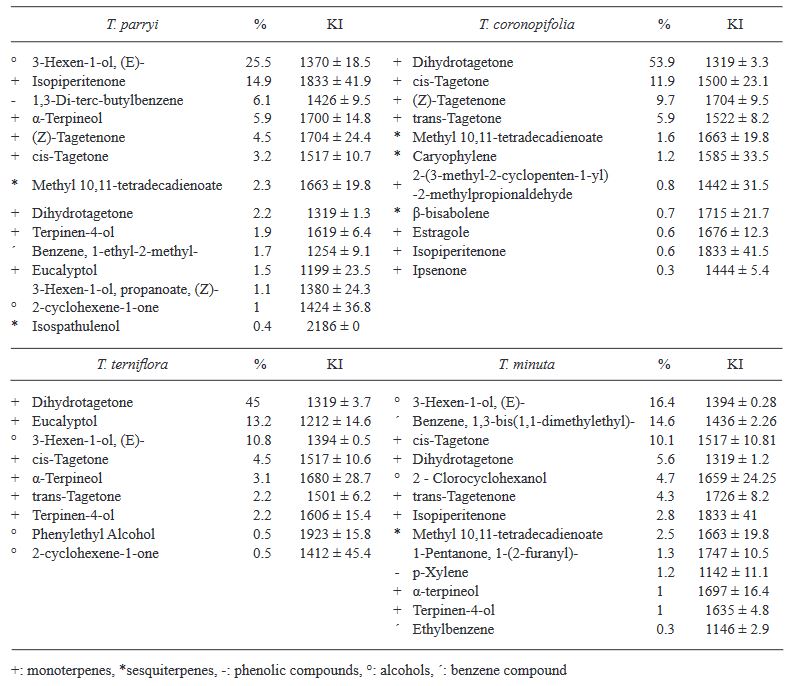
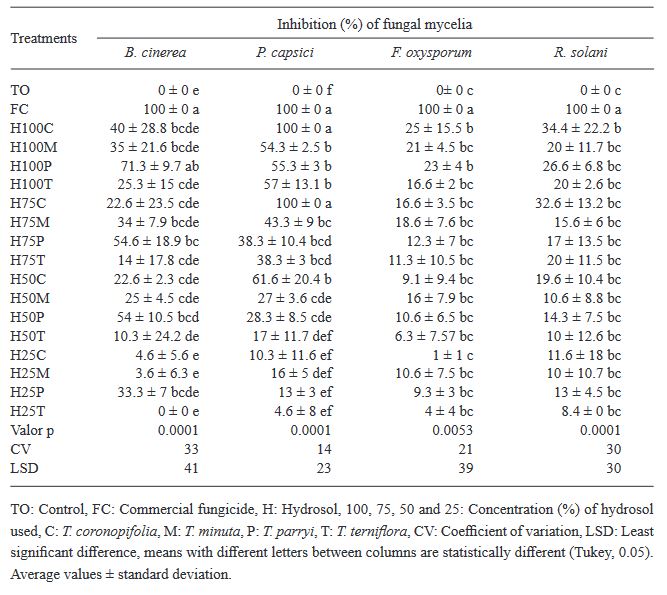
Resumen Antecedentes/Objetivo. Las plantas aromáticas contienen compuestos químicos con potencial para formular productos antifúngicos. El objetivo de esta investigación fue caracterizar la composición química en hidrolatos de especies de Tagetes y evaluar su efecto in vitro e in vivo contra hongos asociados a enfermedades en fresa.
Materiales y Métodos. Los hidrolatos de T. coronopifolia, T. minuta, T. parryi y T. terniflora fueron analizados mediante cromatografía de gases acoplado a detector selectivo de masas. Se evaluaron in vitro hidrolatos al 100, 75, 50 y 25 % y fungicidas comerciales Promyl contra Botrytis cinerea, Fusarium oxysporum, Rhizoctonia solani y Ridomil Gold contra Phytophthora capsici. En la evaluación in vivo las plantas de fresa se asperjaron con hidrolatos y 24 h después fueron inoculadas con suspensión de esporas 1 x 106. Los datos se analizaron mediante análisis de varianza y pruebas de medias de Tukey (p ≤ 0.05).
Resultados. Los monoterpenos fueron los compuestos mayoritarios en las cuatro especies. El hidrolato de T. parryi in vitro inhibió totalmente el crecimiento de B. cinerea y fue efectivo como tratamiento preventivo en la evaluación in vivo. F. oxysporum, P. capsici y R. solani fueron menos susceptibles a todos los hidrolatos.
Conclusión. El hidrolato de T. parryi se puede aplicar como preventivo contra B. cinerea en plantas de fresa
-
O copiar el link
PorZoila Lizbeth Chavarría Cervera, Andrés Quezada Salinas, Pedro Valadez Ramírez, Wilberth Chan Cupul, Jesús Enrique Castrejón Antonio, Juan Carlos Sánchez Rangel*
Recibido: 16/1/2024 – Publicado: 02/4/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2401-2
Resumen Antecedentes/Objetivo. El nopal verdura (Opuntia ficus-indica) es una planta de importancia económica, social y cultural en México, sin embargo, una de las enfermedades recurrentes a esta especie vegetal es la Mancha Negra (MN), la cual se genera por el ataque de diferentes hongos fitopatógenos, de ahí que la identificación de los agentes causales de la MN en los cultivos comerciales de nopal sea una etapa relevante para el manejo agronómico eficiente de la MN. El objetivo de este estudio fue identificar los hongos fitopatógenos causales de la MN en plantaciones de nopal verdura en el estado de Colima, México.
Materiales y Métodos: Se recolectaron 50 cladodios de 50 plantas con síntomas de MN en plantaciones comerciales de Colima, de los hongos aislados se comprobó la patogenicidad de ellos mediante los postulados de Koch, así mismo se identificaron molecularmente aquellos hongos que generaron los síntomas más severos de MN.
Resultados. Se aislaron 35 hongos de plantas sintomáticas de MN, de los cuales 20 presentaron un crecimiento micelial distinto. Solo seis hongos generaron síntomas de MN; siendo tres de ellos los responsables de generar síntomas severos en cladodios: Alternaria alternata, Corynespora cassiicola, and Neoscytalidium dimidiatum.
Conclusión. La MN es generada por diferentes hongos fitopatógenos, pero este es el primer reporte de C. cassiicola y N. dimidiatum como agentes causales de la MN en nopal verdura.
-
O copiar el link


Armillaria gallica asociado a la pudrición de raíz del aguacate en Michoacán
PorJeny Michua Cedillo*, Daniel Téliz Cedillo, Salvador Ochoa Ascencio, María del Pilar Rodríguez, Alejandro Alarcón, Carlos de León, Gerardo Vázquez Marrufo
Recibido: 30/7/2023 – Publicado: 21/3/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2307-7
Resumen Antecedentes/Objetivo. La pudrición de la raíz y muerte de árboles de aguacate asociada a Armillaria es una enfermedad emergente con alto impacto económico en huertos establecidos en áreas previamente forestales de Michoacán. Actualmente se desconoce con precisión las especies asociadas con síntomas típicos de marchites, amarillamiento, producción excesiva de frutos y micelio subcortical en raíz. El objetivo de esta investigación fue caracterizar molecularmente las especies de Armillaria asociadas a la pudrición de la raíz del aguacate.
Materiales y Métodos. Para la caracterización morfológica y molecular, en extracto de malta-agar, se procesaron 60 muestras de raíz de árboles provenientes de tres huertos comerciales con presencia putativa de Armillaria. ADN de aislados purificados se amplificaron por PCR con genes rpb2 y TEF α-1. Las secuencias se alinearon con MAFFT y se realizaron las filogenias con algoritmo de máxima verosimilitud en IQ-TREE.
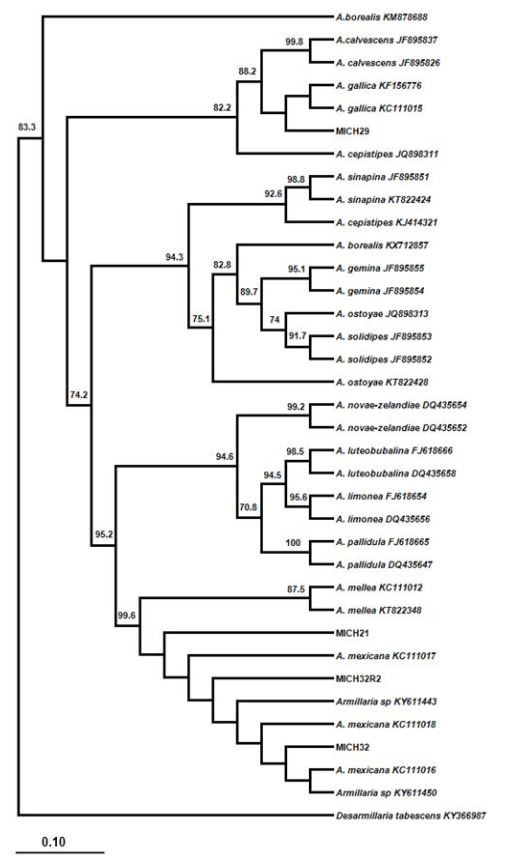
Resultados. Se identificaron dos especies de forma consistente: A. gallica (20%) con 100% de homología y A. mexicana (25%) con 98%. Otra especie que representó el 55% de los aislados no se alineo con ningún grupo. Morfológicamente, los basidiocarpos de A. gallica concuerdan con las características de esta especie. Conclusiones. Este es el primer reporte de A. gallica asociado a la pudrición de la raíz del aguacate en Michoacán.
-
O copiar el link

Recibido: 30/7/2023 – Publicado: 11/4/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2307-3
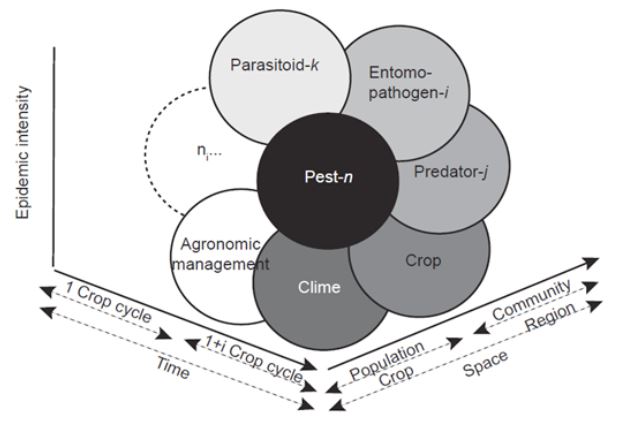
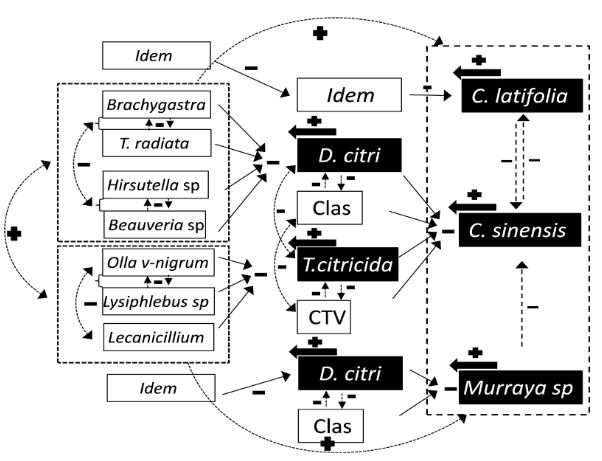
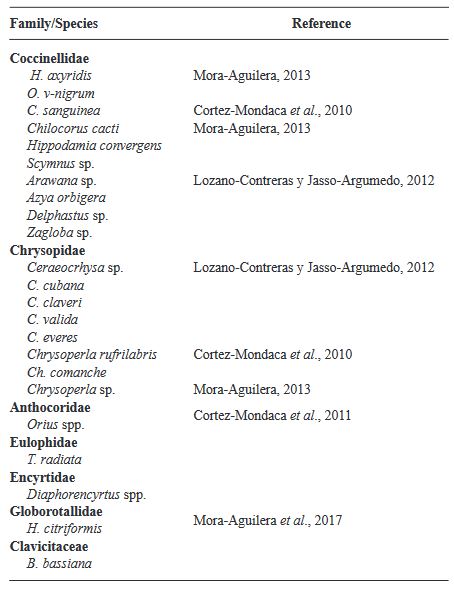
Resumen Diaphorina citri es un importante vector de Candidatus Liberibacter asiaticus (CLas), agente causal del HLB, la enfermedad más devastadora de los cítricos. Aunque se reconoce la importancia del control biológico, los insecticidas son la principal herramienta de control. Sin embargo, su empleo debería ser cauteloso, pues pudiera interferir con el control de plagas reguladas biológicamente. Diaphorina citri cuenta con un amplio abanico de enemigos naturales; sin embargo, solo se aprovecha de forma inundativa al parasitoide Tamarixia radiata y algunas especies de hongos entomopatógenos. Aunque los principales depredadores del vector ocurren de manera natural, pocos estudios abordan su conservación in situ. Esta revisión abona a la idea de que la conservación de enemigos naturales debería ser base del manejo integrado de D. citri y de CLas. Se propone la conservación de hospederos alternos, la inclusión de plantas nectaríferas, conservación de parasitoides in situ, y la autodiseminación de hongos entomopatógenos. Se analizan y discuten estudios sobre conservación de enemigos naturales desarrollados en D. citri y plagas cercanas, su probable impacto en la enfermedad, y perspectivas de implementación en México. Las estrategias propuestas pudieran potenciar, no solo el control biológico de D. citri-CLas, sino también la autorregulación de plagas de cítricos en general
-
O copiar el link


Patosistema de Iris yellow spot orthotospovirus, hospederos del virus y el vector (Thrips tabaci)
PorNorma Ávila Alistac*, Erika J. Zamora Macorra, Héctor Lozoya Saldaña
Recibido: 30/10/2023 – Publicado: 10/3/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-8
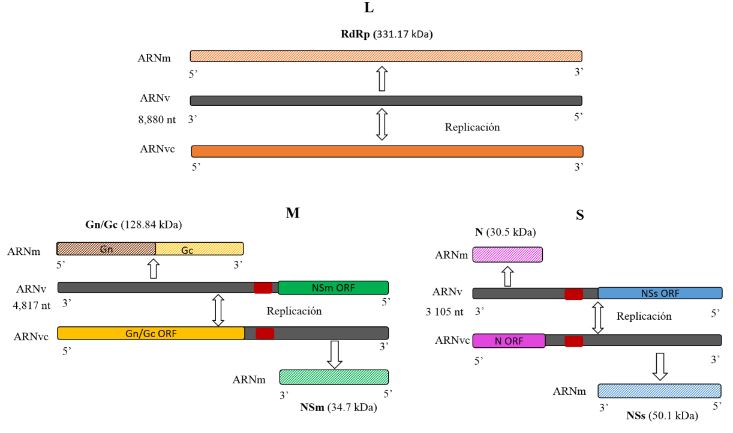
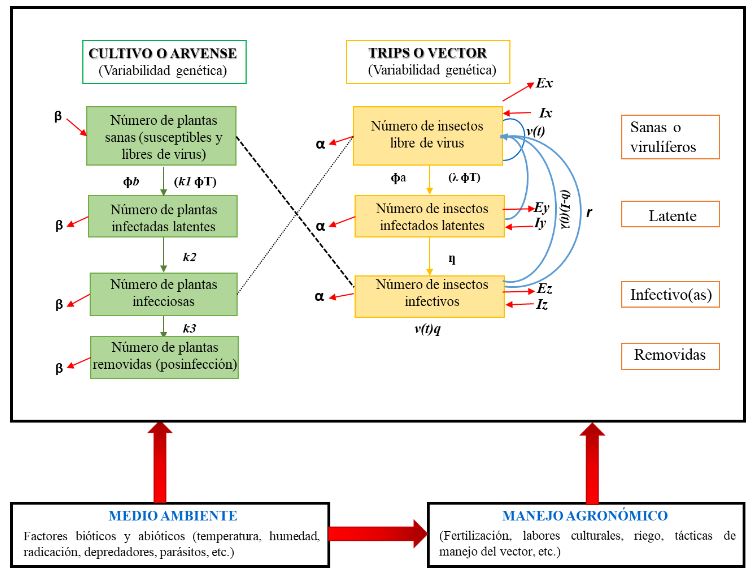
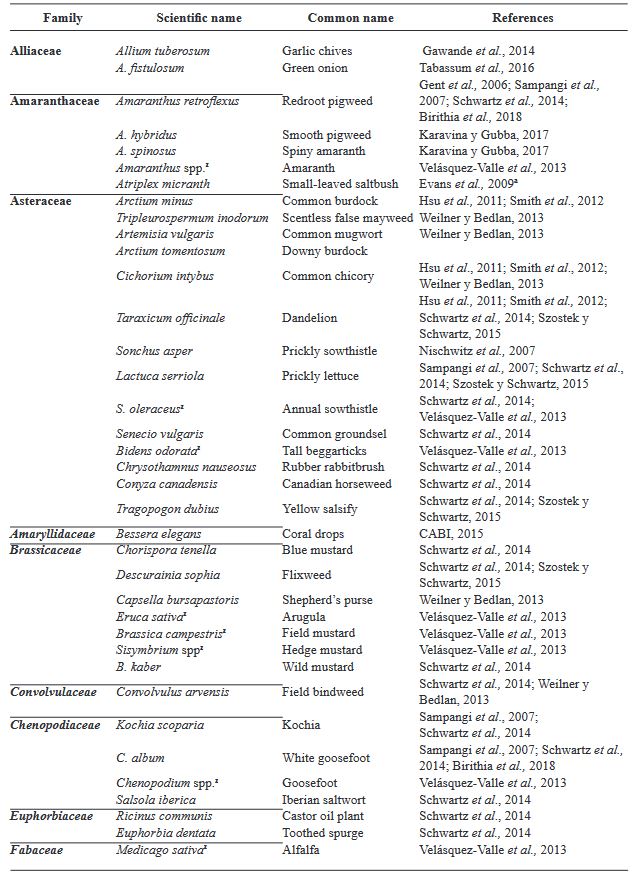
Resumen Iris yellow spot Orthotospovirus (IYSV) causa graves problemas en el cultivo de cebolla (Allium cepa) y está ampliamente distribuido en las zonas productoras del país. En México se reportó en 2010 como “mancha amarilla” en cebolla y en otros miembros del género Allium. Su principal vector Thrips tabaci, que ocasiona daños directos por alimentación y por ser vector de otros virus como Tomato spotted wilt Orthotospovirus e Impatiens necrotic spot Orthotospovirus. El conocimiento del patosistema de IYSV-Thrips tabaci-Allium cepa-arvenses puede coadyuvar a un manejo integral y concientización del uso de pesticidas. La versatilidad de IYSV de infectar a más de 60 especies de plantas (>20 familias), de las cuales en su mayoría están presentes en México; aunado al amplio rango de hospedantes del vector, vuelve compleja la interacción y conlleva a comprender mejor sobre la diversidad de hospedantes alternos del vector y/o el IYSV. En la actualidad, información sobre arvenses como hospederos de IYSV y el vector es limitado, pero su conocimiento proporcionará mayor comprensión de la enfermedad. Es importante tener un conocimiento integral del virus, hospedante principal, hospedantes alternos y vector en el país, para encausar investigaciones futuras para contrarrestar este problema y minimizar pérdidas causadas por IYSV en el cultivo de cebolla principalmente.
-
O copiar el link