
-
O copiar el link


Filtros de Contribuciones
Buscar Contribución:
Control biológico y químico in vitro de hongos asociados a la gomosis en cítricos en Yucatán, México
PorCélida Aurora Hernández Castillo, Patricia Rivas Valencia*, Leticia Robles Yerena, Mariana Guadalupe Sánchez Alonso, Emiliano Loeza Kuk
Recibido: 31/5/2024 – Publicado: 05/11/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-03
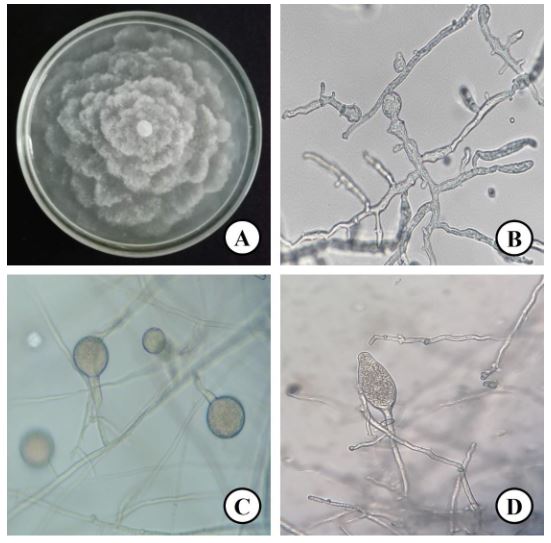
Resumen Antecedentes/Objetivo. En todas las regiones productoras de cítricos en el mundo la gomosis es una enfermedad que ha causado pérdidas en la producción de cítricos. Esta enfermedad es causada por varios patógenos. Los objetivos fueron, identificar los hongos asociados a la gomosis en huertas de cítricos de Plan Chac, Sacalum, Yucatán; y evaluar alternativas químicas y biológicas para el control de los hongos asociados a la gomosis.
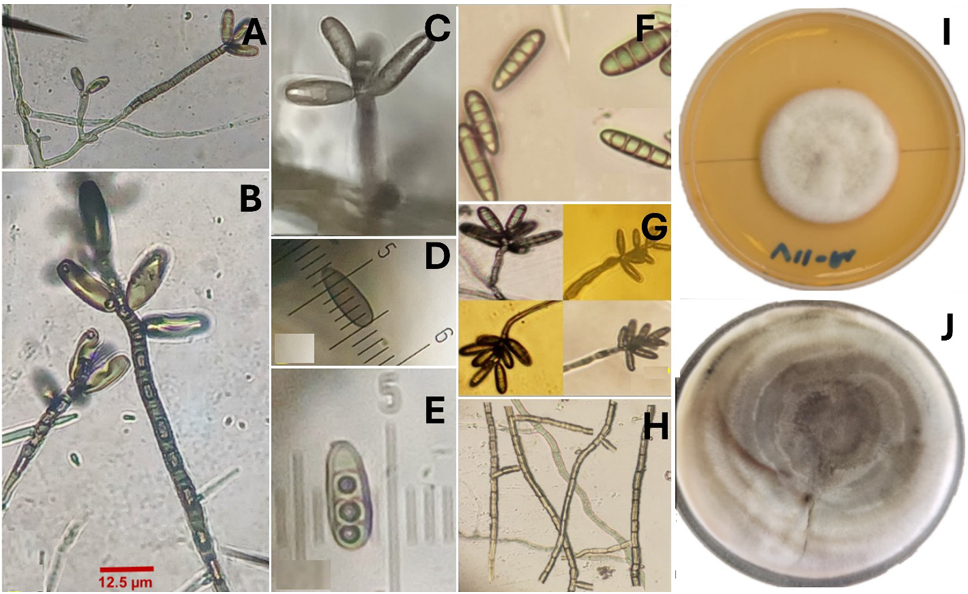
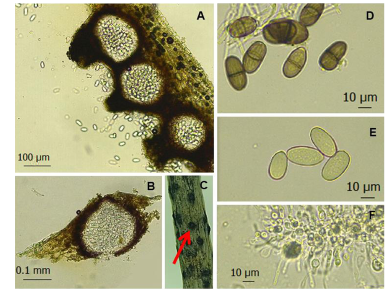
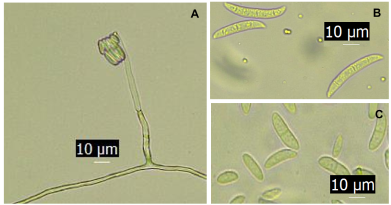
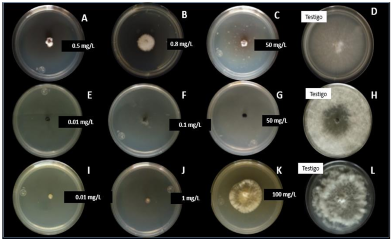
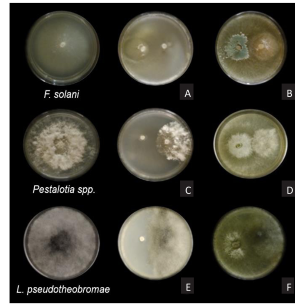
Materiales y Métodos. A partir de tejido vegetal y suelo, se aislaron los hongos asociados. Los aislamientos se identificaron morfológicamente en el tejido vegetal como Lasiodiplodia pseudotheobromae y en suelo como Fusarium solani y Pestalotia spp. La prueba de patogenicidad determinó que L. pseudotheobromae es un agente asociado a ésta enfermedad. Los aislamientos fueron sometidos a pruebas in vitro con fungicidas químicos y agentes antagonistas.
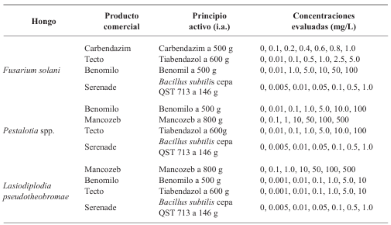
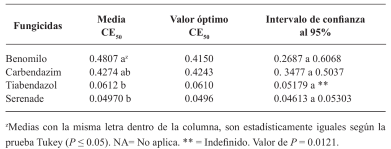
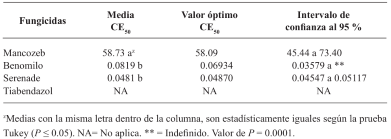
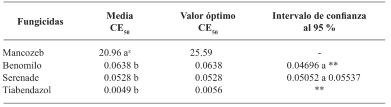
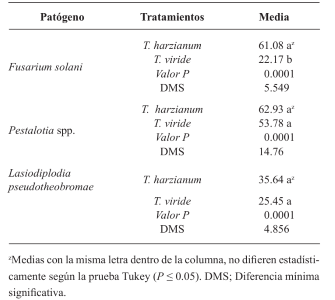
Resultados y Discusión. El Tiabendazol mostró efectividad para F. solani con una concentración efectiva para inhibir el 50 % de la población (CE50) de 0.0612 mg L-1, para Pestalotia spp. se inhibió el crecimiento a todas las concentraciones evaluadas y para L. pseudotheobromae, mostró una CE50 de 0.0049 mg L-1. En el caso de Bacillus subtilis cepa QST 713, disminuyó el crecimiento de F. solani (CE50 0.0496 mg L-1), Pestalotia spp. (CE50 0.0487 mg L-1) y L. pseudotheobromae (CE50 0.0528 mg L-1). Por otro lado, Trichoderma harzianum mostró una mayor inhibición contra F. solani, Pestalotia spp. y L. pseudotheobromae del 61.08, 62.93 y 35.64 %, respectivamente.
Conclusión. En el manejo de gomosis en cítricos se puede incluir con eficiencia el uso de agentes biológicos como Trichoderma y B. subtilis ofreciendo alternativas con menor impacto en el medio ambiente.
-
O copiar el link

PorLeonardo Miguel Nava Eugenio, Dolores Vargas Álvarez, Eleuterio Campos Hernández, Flaviano Godínez Jaimes, Roxana Reyes Ríos, Mairel Valle de la Paz*, Daniel Perales Rosas
Recibido: 31/5/2023 – Publicado: 05/11/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-04
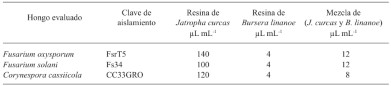
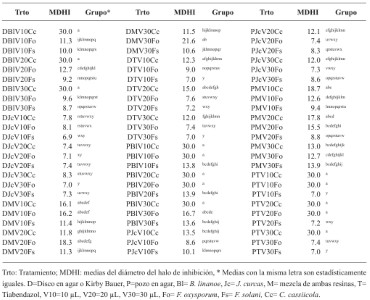
Resumen Antecedentes/Objetivos. Guerrero es importante productor de jamaica (Hibiscus sabdariffa); por lo que el objetivo fue evaluar el efecto inhibitorio de resinas in vitro utilizando cuatro factores: técnica (disco en agar o Kirby Bauer y pozo en agar), resinas (B. linanoe, J. curcas, mezcla de ambas y Tiabendazol), volumen (10, 20 y 30 μL) y hongos fitopatógenos (C. cassiicola, F. oxysporum y F. solani) en el diámetro del halo de inhibición.
Materiales y Métodos. El análisis estadístico se realizó con un diseño factorial completamente al azar de efectos fijos para comparar los 72 tratamientos se usó la prueba de Kruskal-Walis.
Resultados. Se encontró que todos los términos fueron significativos, los efectos principales de técnica, resinas, volumen y hongos en el diámetro del halo de inhibición, pero también las interacciones dobles, triples y cuádruples. .
Conclusión. La resina de B. linanoe mostró mayor inhibición para C. cassiicola y F. oxysporum, en las dos técnicas (técnica de pozo en agar y disco en agar o técnica Kirby Bauer), esto la convierte en el tipo de resina con mayor potencial biocontrolador.
-
O copiar el link
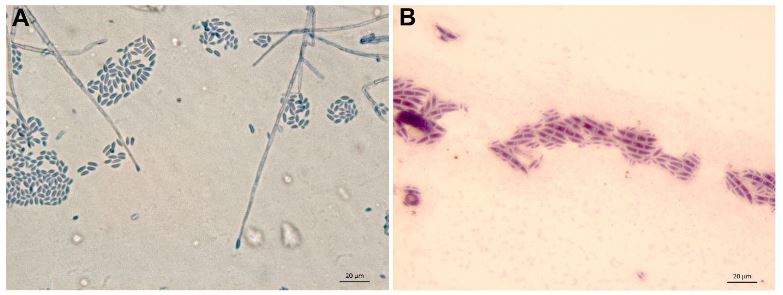
Diversidad y taxonomía de Fusarium solani aislado de plantas marchitas de Agave tequilana var azul
PorViviana Montaño Becerrra, Norma Alejandra Mancilla Margalli, Cristina Chávez Sánchez, Martin Eduardo Avila Miranda*
Recibido: 30/11/2023 – Publicado: 25/10/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-5
Resumen Antecedentes/Objetivo. El objetivo del trabajo fue identificar 24 cepas de F. solani aisladas de agave con marchitez, respecto a las nuevas especies filogenéticas; determinar su semejanza molecular con f. spp. de F. solani; determinar su diversidad genética y su capacidad patogénica en agave, frijol (Phaseolus vulgaris) y maíz (Zea mays).
Materiales y Métodos. Secuencias del fragmento ITS1-5.8S-ITS2 de 24 aislados de agave y de las f. spp. de F. solani, se compararon con el GenBank y FUSAROIDID. Secuencias amplificadas del 18S rRNA, se alinearon con secuencias reportadas de F. solani f. spp. phaseoli y batatas, definiendo presencia de intrones. Se determinó diversidad genética con el marcador de DNA RepPCR. Cepas representativas se confrontaron con plántulas de agave, frijol y maíz, evaluando su patogenicidad como severidad de pudrición radicular.
Resultados. Aislados morfológicamente identificados como F. solani, el GenBank los ubicó como F. solani o incluidos en el FSSC, tres cepas se identificaron como Xenoacremonium sp. FUSAROID-ID definió que las secuencias de F. solani eran altamente similares a las de Neocosmospora martii, N. pseudoradicicola, N. solani y N. falciformis. Las secuencias ITS1-5.8S-ITS2 y ausencia de intrones en su SSU, indicó que ninguno es F. solani f. sp. phaseoli. Aislados obtenidos de agave fueron patogénicos a A. tequilana y a un cv de maíz criollo, pero no a maíces con resistencia a Fusarium. Ningún aislado de agave fue patogénico a frijol.
Conclusiones. Cuatro especies filogenéticas del FSSC provocan pudrición radicular en agave; aislados de F. solani de agave no afectaron a maíces resistentes a Fusarium. Es seguro intercalar frijol en el cultivo de agave.
-
O copiar el link

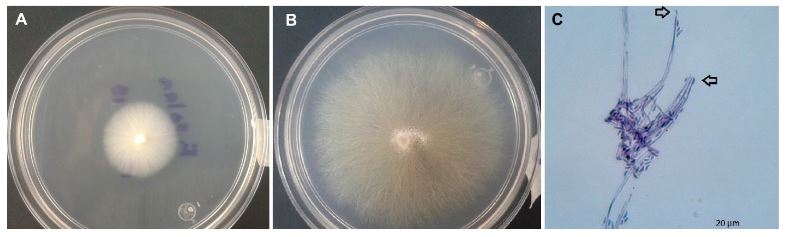
PorDiana Angelina Urias Lugo, Octavio Ernesto Martínez Ereva, Cecilia de Los Ángeles Romero Urías, Carlos Ramiro Ibarra Sarmiento, Sylvia Adriana Estrada Díaz, Rubén Félix Gastélum, Karla Yeriana Leyva Madrigal, Guadalupe Arlene Mora Romero*
Recibido: 24/5/2024 – Publicado: 24/10/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-01
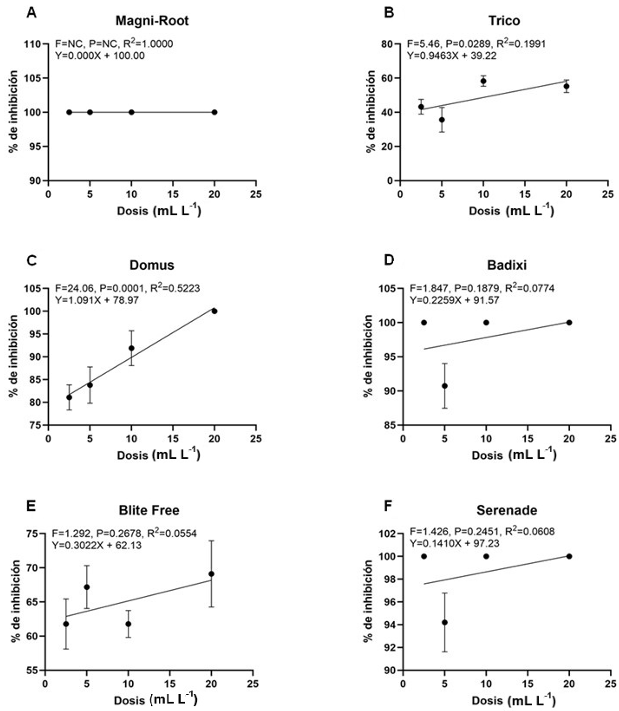
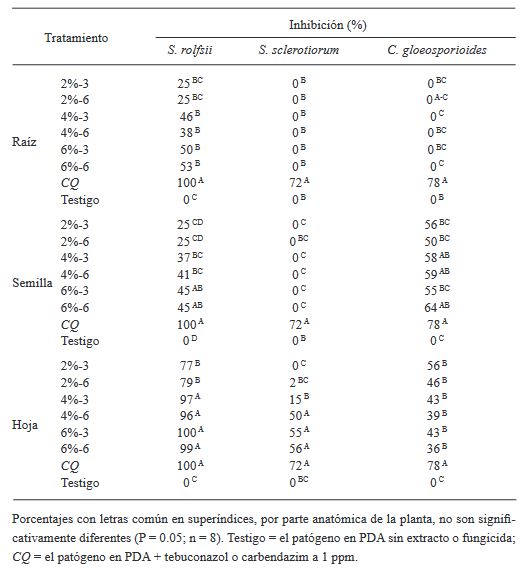
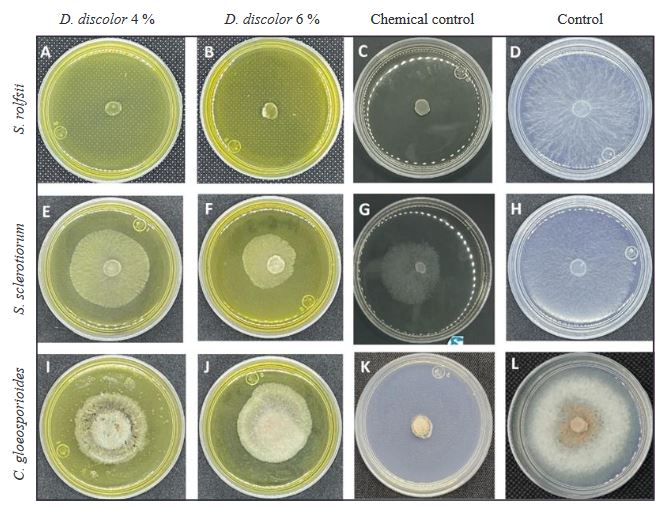
Resumen Antecedentes/Objetivo. El presente trabajo reporta el efecto in vitro de extractos acuosos (2, 4 y 6% p/v) de raíz, semilla y hoja de Datura discolor obtenidos en dos tiempos (3 y 6 minutos) por procesamiento de alta presión, contra Sclerotium rolfsii, Sclerotinia sclerotiorum y Colletotrichum gloeosporioides.
Materiales y Métodos. Se prepararon extractos de raíces, semillas y hojas de D. discolor en una proporción de 1:10 p/v con agua destilada. Se consideraron dos tratamientos continuos de alta presión (600 MPa) manteniendo la presión durante 3 min y otro con presión (600 MPa) mantenida durante 6 min. Los extractos se evaluaron contra S. rolfsii, S. sclerotiorum y C. gloeosporioides. Los experimentos se realizaron en cajas Petri con medio PDA. Se evaluó la eficiencia de los extractos obteniendo el porcentaje de inhibición.
Resultados. Los resultados mostraron porcentajes variables de inhibición de los extractos en las distintas partes de la planta y concentraciones; los extractos de hoja al 6%, independientemente del tiempo de extracción mostraron efectividad contra los tres patógenos, con inhibición de 99 y 100%, de 55 y 56%, y de 43 y 36%, para S. rolfsii, S. sclerotiorum y C. gloeosporioides, a los 3 y 6 minutos respectivamente.
Conclusión. La efectividad del extracto de hoja al 6%, seis meses posteriores a la preparación fue similar al de los extractos en fresco. Estos resultados abren a futuras líneas de investigación orientadas hacia el manejo sustentable de fitopatógenos. Se sugieren estudios sobre efectividad biológica de los extractos en invernadero y campo.
-
O copiar el link
Tobamovirus fructirugosum una enfermedad emergente: revisión y situación actual en México
PorUbilfrido Vásquez Gutiérrez, Juan Carlos Delgado Ortiz*, Gustavo Alberto Frías Treviño, Luis Alberto Aguirre Uribe, Alberto Flores Olivas
Recibido: 28/1/2024 – Publicado: 15/10/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2401-7
Resumen Antecedentes/Objetivo. La especie Tobamovirus fructirugosum (ToBRFV) es considerado una plaga cuarentenaria a nivel mundial que limita la producción de Solanum lycopersicum y Capsicum annum, actualmente presente en tres países del continente americano. El objetivo de este trabajo fue profundizar en la variabilidad genética del ToBRFV con respecto a los diversos aislados, la caracterización físico-molecular y sintomática, los métodos tradicionales y más actuales implementadas para el diagnóstico, rango de hospedantes reservorios del virus, y la epidemiología. Resultados. ToBRFV se generó de una mutación resultado de la recombinación genética con TMV, considerado principal progenitor y ToMMV progenitor secun¬dario. Análisis filogenéticos reportan la existencia de cinco clados con respecto a la diversidad genética del ToBRFV. Los primeros cebadores para la detección se diseñaron en 2015 que codifican proteínas de replicación, movimiento y cápside. Los métodos serológicos pueden ser utilizados para un diagnóstico preventivo, mientras que las moleculares y NGS pueden confirmar la infección por el virus aún en bajas concentraciones en la planta. Se reportan 16 familias de malezas y cultivos hospedantes, registrados en 47 países. Para lograr una estrategia efectiva, es necesario disminuir las fuentes de inóculo, desarrollar compuestos inhibidores de la transmisión mecánica y el desarrollo de genotipos tolerantes. Conclusión. ToBRFV está distribuido a nivel nacional, y representa un riesgo fitosanitario para México; el análisis exhaustivo del estudio de técnicas de diagnóstico, rango de hospedantes, diseminación, epidemiología y estrategias de control, contribuye al conocimiento del ToBRFV.
-
O copiar el link



PorMaria Magdalena Rivera Salas, José Basilio Heredia, Juan Manuel Tovar Pedraza, Cesar San Martín Hernández, José Benigno Valdez Torres, Isabel Cruz Lachica, Raymundo Saúl García Estrada*
Recibido: 22/5/2024 – Publicado: 23/8/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2405-11
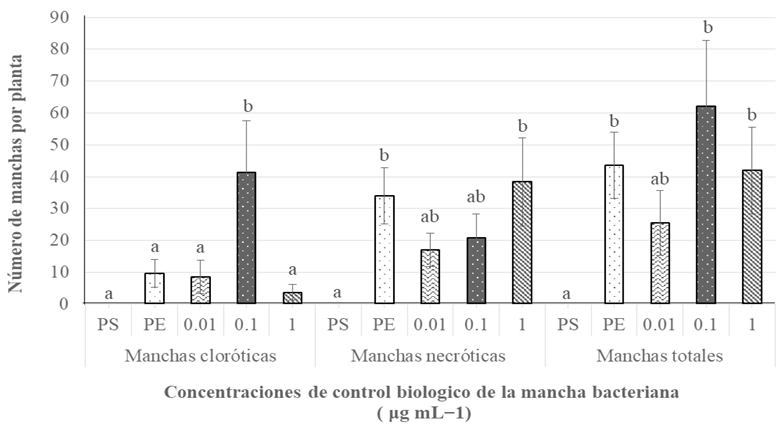
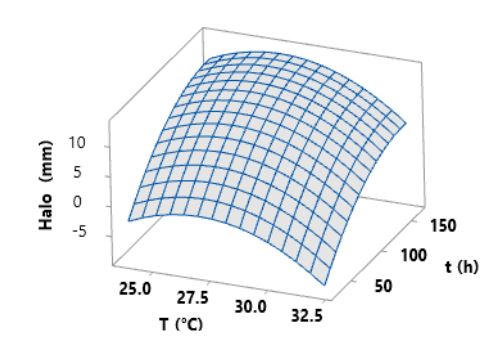
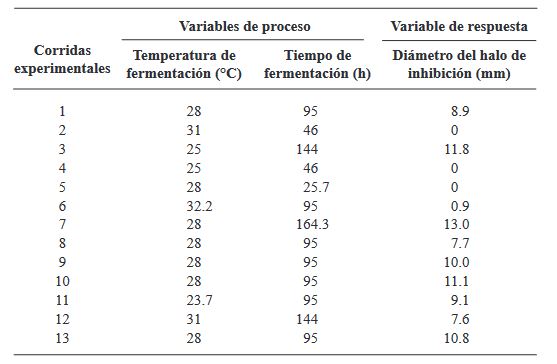
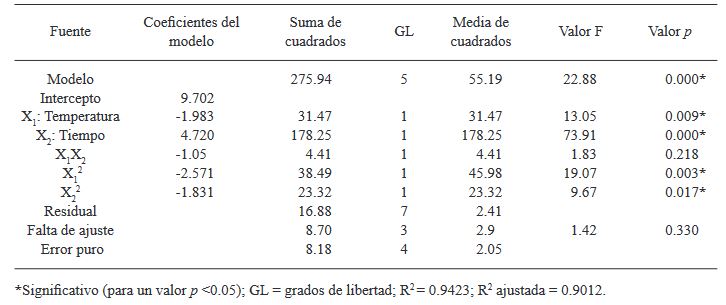
Resumen Antecedentes/Objetivo. Actualmente, las especies del género Bacillus están ganando interés debido a su capacidad para producir metabolitos secundarios con propiedades antifúngicas contra diversos hongos fitopatógenos. El objetivo de este estudio fue optimizar la temperatura y el tiempo de fermentación para la producción de un extracto antifúngico de Bacillus amyloliquefaciens B17 y verificar su actividad contra Gilbertella persicaria, Choanephora cucurbitarum, Colletotrichum asianum y Botrytis cinerea.
Materiales y Métodos. Se utilizó un diseño central compuesto (DCC) con dos factores y cinco niveles (temperatura de fermentación: 23.7, 25, 28, 31 y 32.2 °C y tiempo de fermentación: 25, 46, 95, 144 y 164.3 h). Trece combinaciones de temperatura y tiempo de fermentación se realizaron de manera aleatoria. Los trece extractos crudos de B. amyloliquefaciens B17 se obtuvieron, del caldo de fermentación libre de células, mediante precipitación ácida seguida de solubilización alcalina. La variable de respuesta fue el diámetro de los halos de inhibición generados al colocar gotas de los diferentes extractos crudos sobre el medio inoculado con una suspensión de esporas de Gilbertella persicaria.
Resultados. Las condiciones óptimas para la producción del extracto con mayor actividad antifúngica en B. amyloliquefaciens B17 fueron 26.8 °C y 158.6 h.
Conclusión. El extracto crudo optimizado de B. amyloliquefaciens B17 exhibió una gran capacidad para inhibir el crecimiento micelial y la germinación de esporas de Gilbertella persicaria, Choanephora cucurbitarum, Colletotrichum asianum y Botrytis cinerea.
-
O copiar el link
PorErick Ortega Piña, Daniel Leobardo Ochoa Martínez*, Reyna Isabel Rojas Martínez, Alfredo Díaz Lara
Recibido: 15/2/2024 – Publicado: 12/8/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2402-8
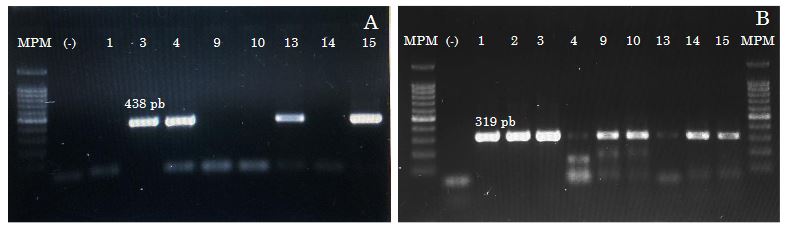
Resumen Antecedentes/Objetivo. En cultivos de haba (Vicia faba) establecidos en Montecillo, Texcoco, Estado de México se observó un 100 % de plantas con síntomas de virosis consistentes en mosaico, moteado, enrollamiento de hojas, marchitamiento y un decremento notable en el desarrollo de las plantas. Estos síntomas condujeron a una reducción significativa en el rendimiento y calidad de las semillas. En este contexto, se realizó el presente estudio con el objetivo de identificar las especies virales asociadas con estos síntomas.
Materiales y Métodos. La estrategia experimental consistió en extracción de ARN total de hojas manifestando los síntomas mencionados, seguido de secuenciación de nueva generación.
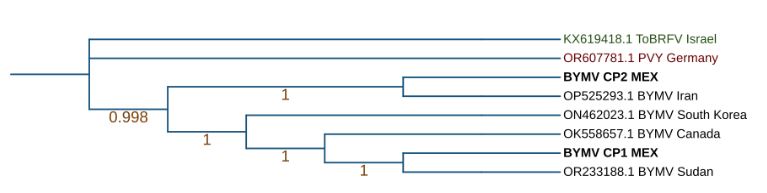
Resultados. En los resultados se obtuvieron los genomas completos de Orthotospovirus impatiensnecromaculae (antes impatiens necrotic spot virus) y de dos aislamientos del Potyvirus phaseoluteum (antes bean yellow mosaic virus). El análisis filogenético reveló que el aislamiento de O. impatiensnecromaculae (INSV CPMEX) muestra divergencias significativas respecto a los previamente reportados en otras especies vegetales. Por otro lado, los dos aislamientos de P. phaseoluteum (BYMV CP1MEX y BYMV CP2MEX) demostraron ser distintos entre sí, estando relacionados con aislamientos reportados en Sudán e Irán.
Conclusión. Este hallazgo sugiere una diversidad genética considerable entre los virus asociados a los síntomas de virosis en cultivos de haba en la región, lo que subraya la importancia de una identificación precisa para el manejo y control de estas infecciones virales
-
O copiar el link

PorYeison David López Galé*, Mauricio Fernando Martínez, Lizeth Paola Palacios Joya, Nubia Murcia Riaño, Mario Augusto García Dávila
Recibido: 22/11/2023 – Publicado: 08/8/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2311-1
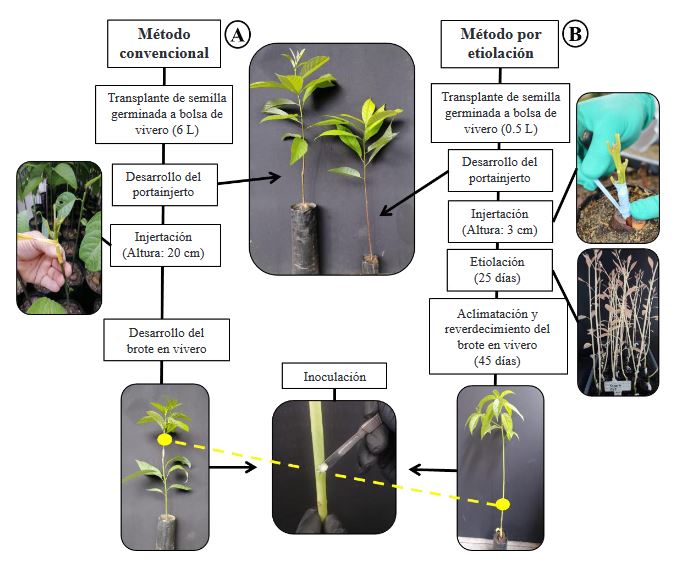
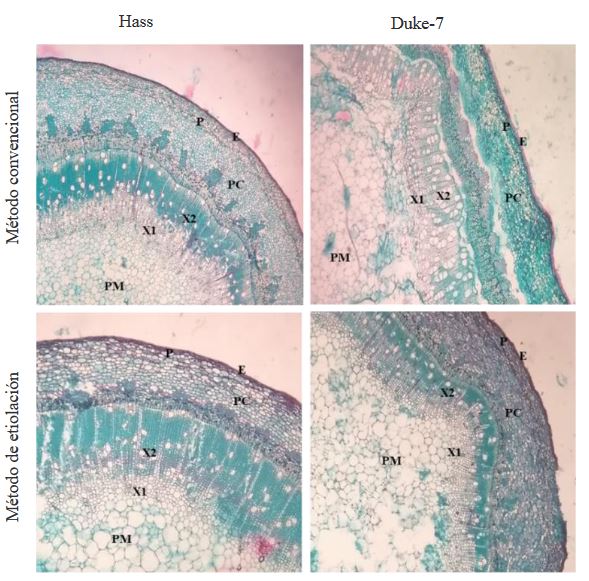
Resumen Antecedentes/Objetivo. El nivel de resistencia a Phytophthora cinnamomi en germoplasma de aguacate puede ser evaluado de forma indirecta a través de la inoculación del patógeno por herida al tallo. El objetivo de este trabajo fue comparar el método de desarrollo convencional de injertos y el método de injertos etiolados para determinar niveles de resistencia indirecta a P. cinnamomi a través de la técnica de inoculación por herida al tallo.
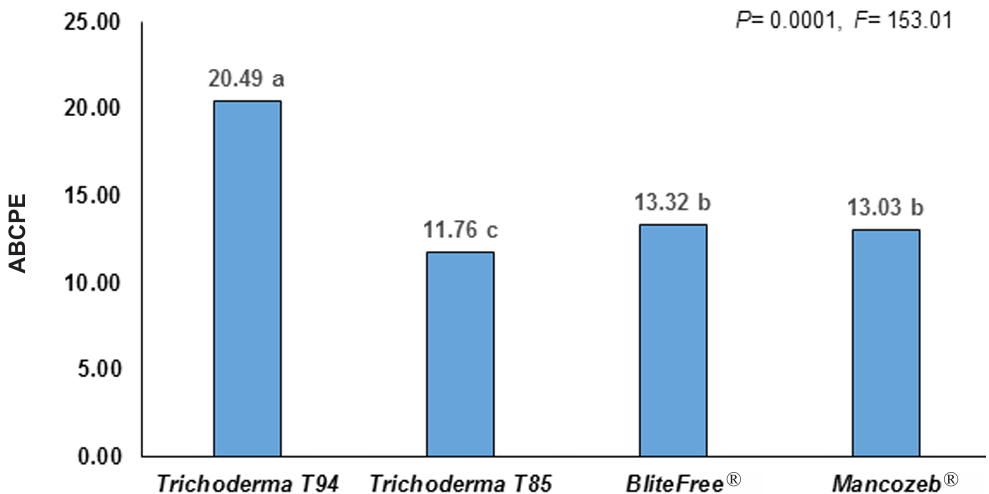
Materiales y Métodos. En el estudio, se utilizaron tres aislamientos de P. cinnamomi y dos genotipos de aguacate con diferente nivel de resistencia al pató geno, Duke-7 (medianamente resistente) y Hass (susceptible). La multiplicación clonal de los genotipos se realizó con yemas injertadas sobre portainjertos propagados por semillas de aguacate antillano. La inoculación fue realizada en el brote a la altura de 8 cm y el crecimiento de las lesiones se midieron durante 24 días. Con los datos, se determinó el Área Bajo la Curva del Progreso de la Enferme dad (ABCPE) y el Coeficientes de Variación (CV). La información fue analiza da con un diseño experimental completamente al azar con arreglo factorial 2*2*3 (Método*Genotipo*Aislamiento).
Resultados. El análisis de varianza para el ABCPE no mostró diferencias entre métodos (p=0.1881); sin embargo, se presentaron diferencias entre genotipos, aislamientos y entre las interacciones genotipo*método y genotipo*aislamiento (p≤0.05). Con el método convencional, el desarrollo de los brotes fue tardío (141- 159 días) y el tamaño de las lesiones fue altamente variable (CV=38.9-64.4 %). Los brotes etiolados y reverdecidos en vivero, por el contrario, presentaron rápido crecimiento (101-107 días) y mayor uniformidad en las lesiones generadas por el patógeno (CV=11.1-24.2 %), lo que permitió discriminar niveles de resistencia entre genotipos, así como grados de agresividad entre aislamientos.
Conclusión. El desarrollo de brotes etiolados en injertos de aguacate se propone como un método alternativo rápido que puede garantizar mayor uniformidad en el desarrollo de lesiones dentro de las unidades experimentales de un tratamiento, logrando de esta manera mayor confiabilidad al momento de evaluar y seleccionar de forma preliminar genotipos de aguacate con atributos de resistencia indirecta a P. cinnamomi.
-
O copiar el link
Infecciones virales mixtas en cultivos de hortalizas: aspectos bioquímicos y moleculares
PorMario Sánchez Sánchez, Irasema Vargas Arispuro, Juan Manuel Tovar Pedraza, Cristóbal González Pérez Pedraza, Emmanuel Aispuro Hernández, Eber Addí Quintana Obregón, Miguel Ángel Martínez Téllez*
Recibido: 05/4/2024 – Publicado: 06/8/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-3
Resumen Las infecciones virales mixtas se refieren a la coinfección de dos o más virus en la planta, que regularmente provocan síntomas exacerbados en hojas y frutos. La dinámica de las coinfecciones puede seguir una interacción sinérgica, antagónica o neutral que afecta la gravedad de los síntomas y la infección. Las infecciones virales mixtas ocurren debido a la convergencia de características fundamentales revisadas en este manuscrito. La interrelación virus-planta huésped influye en el establecimiento y los patrones de propagación de infecciones virales mixtas. Se debe prestar atención a los posibles cambios en la dinámica de transmisión y prevalencia de enfermedades virales de las plantas debido al efecto de alteraciones antropogénicas y naturales en sistemas agroecológicos complejos o sus componentes, incluidos huéspedes, reservorios, vectores, nichos ecológicos y la aparición de nuevas cepas de virus.
-
O copiar el link
![Figura 1. Transición de síntomas en las hojas inferiores, medias y superiores de plantas de tomate agroinoculadas con los componentes de ADN-A y DNA-B de clones infecciosos de aislados de TYLCV (ToYMoV-[CR:Gre:GR1:90]), y Tomato leaf curl Sinaloa virus (ToLCSiV-[CR:Lib:L1:02]) de Costa Rica y un clon infeccioso del ADN genó mico de TYLCV de la República Dominicana (TYLCV-[DO]) de forma individual o en todas las combinacio nes. (A) TYLCV; (B) planta de tomate no inoculada; (C) ToYMoV y ToLCSiV; (D) ToYMoV y TYLCV; (E) ToLCSiV y TYLCV; (F) ToYMoV, ToLCSiV y TYLCV. Las plantas fueron fotografiadas 21 días después de la agroinoculación (con el permiso de Maliano <em>et al.,</em> 2022)](img/RMF/Volumenes/NumNormales/V4232024/RMF2404-3/Figure1.jpg)
PorRubén Félix Gastélum*, Gabriel Herrera Rodríguez, Norma Ávila Alistac, Elizabeth León
Recibido: 22/4/2024 – Publicado: 06/8/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-6
Resumen El moho blanco (Sclerotinia sclerotiorum) es la principal enfermedad del frijol y de la papa en Sinaloa. Se describen los síntomas y signos de la enfermedad, las características culturales y morfológicas del teleomorfo del patógeno, su ecología y la epidemiología de la enfermedad. Se aborda la implementación de un sistema de predicción que contempla la germinación carpogénica de los esclerocios y la fenología del frijol y de la papa para el manejo de la enfermedad. Dicho sistema incluye la temperatura del suelo a 2.5 cm de profundidad y la floración en dichos cultivos. Una vez que en el suelo ocurren temperaturas de 13 a 19 °C e inicia la floración en los cultivos, se realiza la primera aplicación preventiva de fungicida sintético contra la enfermedad. Estudios de laboratorio indicaron que Trichoderma harzianum, T. viride y T. atroviride ejercen inhibición in vitro contra S. sclerotiorum y controlaron el moho blanco en campo, pues hubo un incremento en el rendimiento de 40 %, con respecto a las parcelas que se aplicaron con el fungicida fluazinam. Se proponen nuevas líneas de investigación enfocadas a la ecología del patógeno y al manejo de la enfermedad donde se incluyan estos hongos benéficos en el sistema de predicción.
-
O copiar el link
