Share this link via
Or copy link


Article filters
Search Papers
byAbimael Rubio Sosa, Misael Martínez Bolaños, Juan Florencio Gómez Leyva, Salvador Lozano Trejo*, Ernesto Castañeda Hidalgo, Gustavo Omar Diaz Zorrilla
Received: 30/August/2024 – Published: 26/March/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2307-1
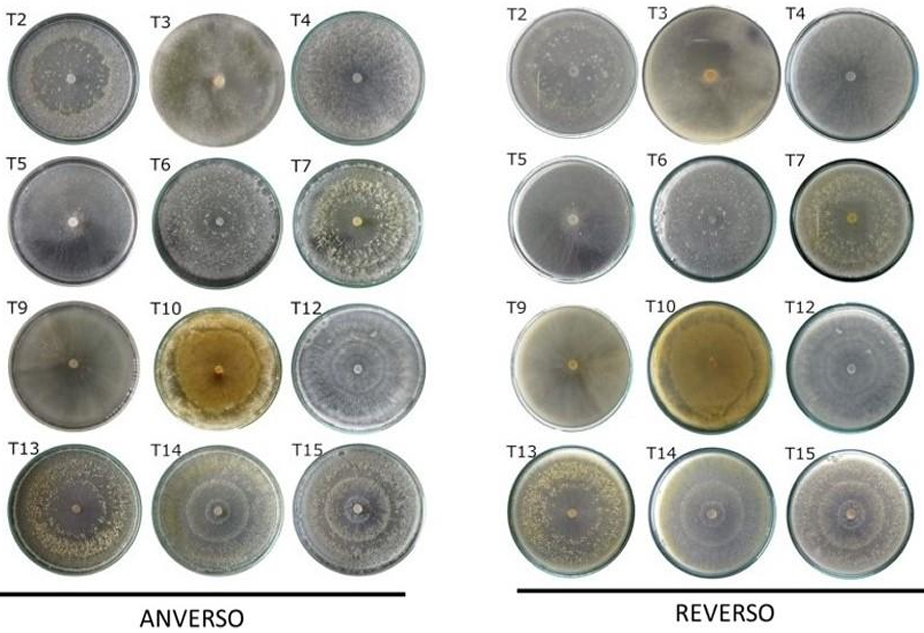
Abstract Background/Objective. The objective of the study was to isolate and characterize native isolate of Trichoderma from organic crops of Arabica coffee (Coffea arabica) in Oaxaca state, as well as to evaluate their in vitro biocontrol potential against Colletotrichum spp., causal agent of anthracnose.
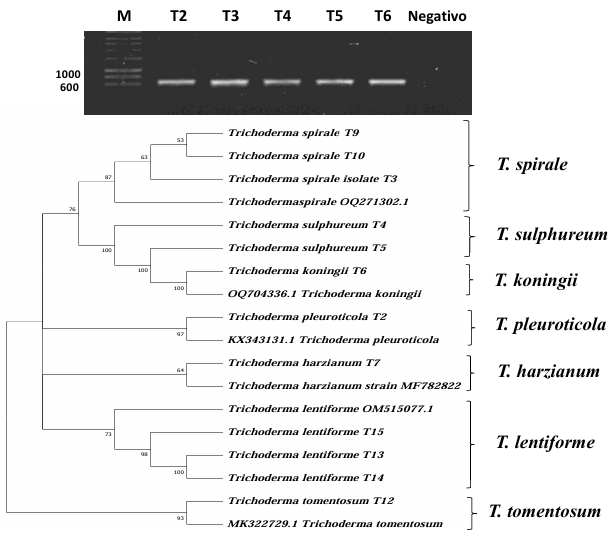
Materials y Methods. Soil and vegetative material samples were collected from coffee plant plots, from which fungal strains corresponding to the genera Trichoderma and Colletotrichum were isolated. Macroscopic and microscopic characterization was performed and the growth rate of each of the isolates was evaluated. Finally, molecular characterization was performed by sequencing the ITS region of rRNA. To evaluate the biocontrol potential, antagonism tests were performed between the isolates of the two genera.
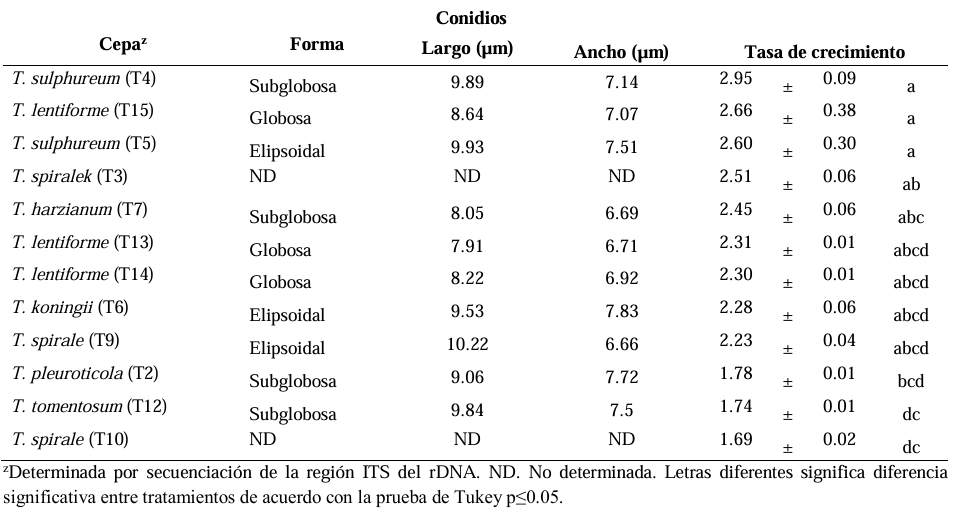
Results. Seven different species were identified: T. harzianum, T. pleuroticola, T. sulphureum, T. tomentosum, T. koningii, T. spirale and T. lentiforme. The latter were the most abundant. Of these, T. lentiforme was selected and evaluated for its in vitro inhibition capacity against three Colletotrichum spp. It was observed that the growth of the fungus was inhibited by 20 to 80%.
Conclusion. The potential of Trichoderma as a biocontrol agent for Colletotrichum spp. is highlighted, acting in different ways against this phytopathogen. This contributes to the knowledge about the diversity of native Trichoderma species, to the coffee-growing region of the state of Oaxaca. In addition, this deeper knowledge contributes to enriching knowledge and choosing these species for future studies in the biocontrol of phytopathogens, in order to promote sustainable agricultural practices.
Share this link via
Or copy link


byTabita Queren Pérez Reyes, Santos Gerardo Leyva Mir, Mario Pérez Grajales*, María Teresa Martínez Damián, Rogelio Castro Brindis
Received: 30/August/2024 – Published: 08/March/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2408-5
Abstract Background/Objective. Phytophthora capsici causes losses of up to 100 % in Capsicum pubescens and there are no resistant commercial varieties. A viable and sustainable alternative is to use the CM-334 rootstock (Capsicum annuum), which is universally resistant to Phytophthora capsici.
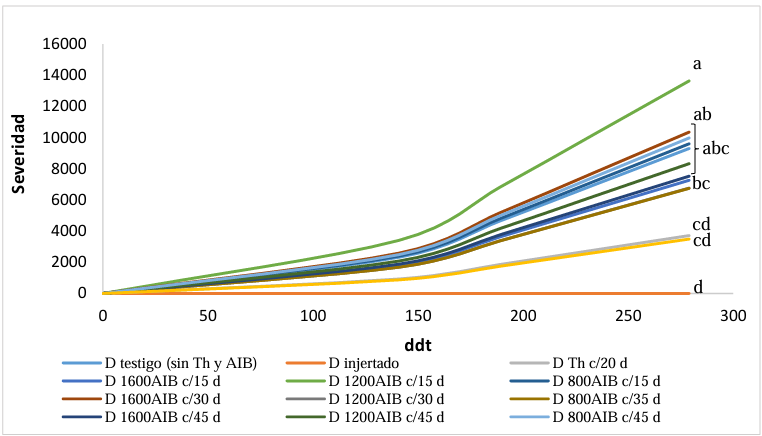
Materials and Methods. The following was studied: the root biomass of CM-334 when grafting the mazano chili hybrids ‘Maruca’, ‘Jhos’, and ‘Dali’, the resistance of the graft to P. capsici in infested soil and its yield (hybrid ‘Dali’), and the root biomass of CM-334 with applications of auxins and T. harzianum.
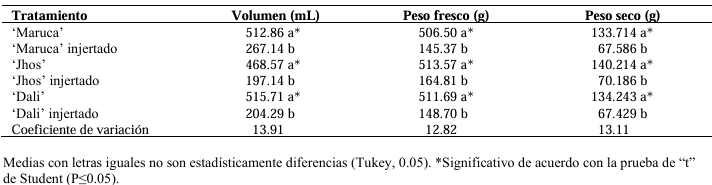
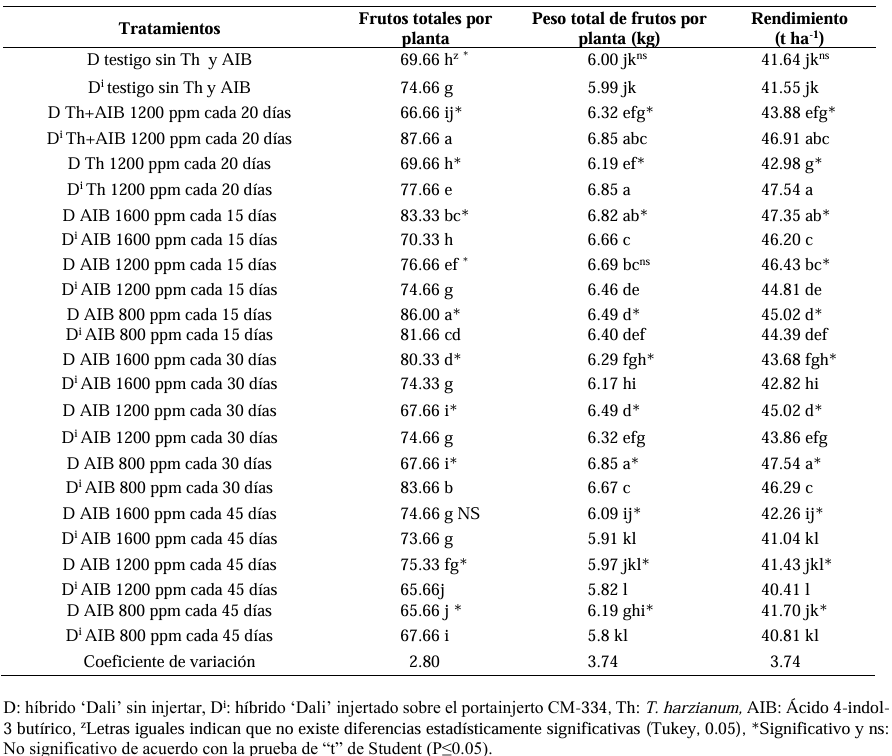
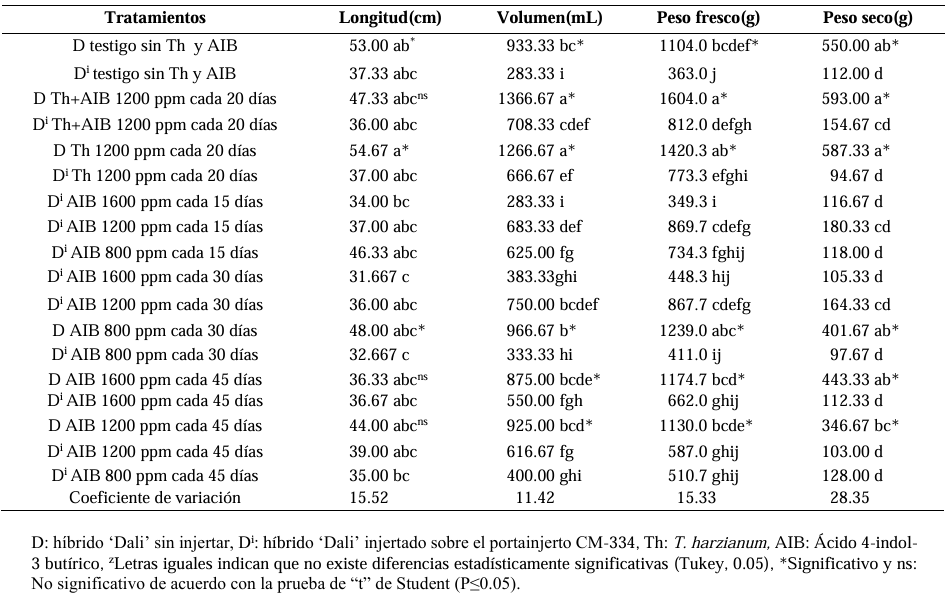
Results. As a rootstock, CM-334 exhibited 50, 53 and 75 % less root volume, fresh weight, and dry weight, respectively, compared to non-grafted hybrids. Using the CM-334 rootstock, there was no incidence of P. capsici and the yield decreased by 2 %, and even with T. harzianum, alone or in combination with 1200 ppm of IBA, the yield increased by 8 %. The grafted ‘Dali’ hybrid had 32, 50, 50, and 76 % less root length, volume, fresh weight, and dry weight, respectively, compared to the non-grafted hybrid; therefore, it is suggested to apply 1.25 kg ha-1 of T. harzianum and 1200 ppm of IBA every 20 days to improve root biomass.
Conclusion. Grafting manzano chili onto CM-334 is a viable and sustainable control alternative to reduce P. capsici incidence since none of the grafted plants showed wilt symptoms like the non-grafted ones, and the yield was the same as in the first production cycle, with the advantage that grafted plants produce more cycles (4 years), whereas the non-grafted ones die during the first cycle because of the oomycete.
Share this link via
Or copy link


byGermán Ramírez Jiménez, Omar G. Alvarado Gómez, Magdiel Torres de la Cruz*, Miguel Ángel Mayo Hernández, Ángel F. Huamán Pilco, Jorge R. Díaz Valderrama
Received: 07/June/2024 – Published: 21/February/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2406-2
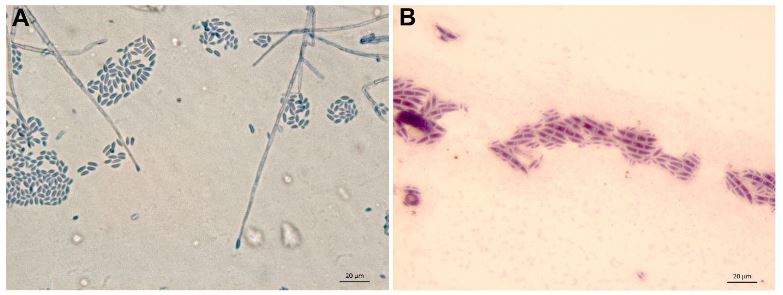
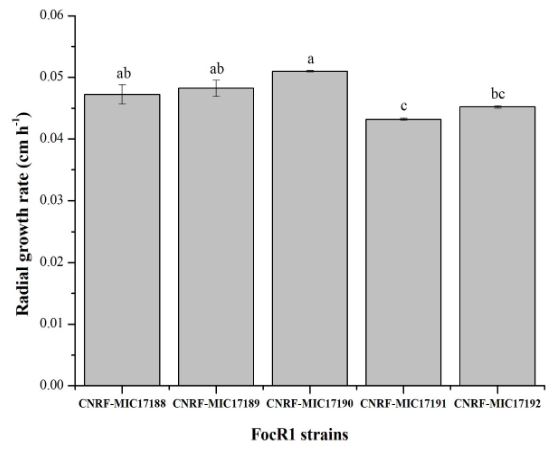
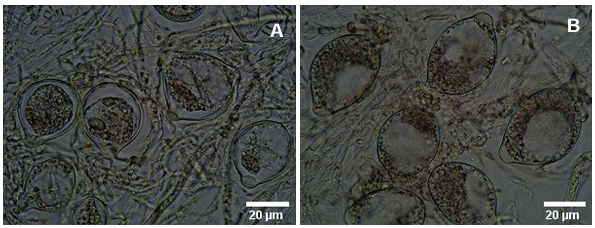
Abstract Background/Objective. Garlic (Allium sativum) is a crop of economic relevance in Mexico. Nuevo León stands out in production; however, in the municipality of Aramberri, post-harvest losses have been reported due to diseases of unknown etiology. The objective of this work was to identify the fungi associated with the postharvest deterioration of A. sativum bulbs in Aramberri, Nuevo León, Mexico and to evaluate their in vitro sensitivity to fungicides.
Materials and Methods. From bulbs with evidence of deterioration and necrosis, fungi were isolated in PDA medium. Four isolates were identified by morphological analysis and one isolate from each morphological species was identified by molecular analysis. The pathogenicity of the four isolates on symptom-free bulbil was evaluated. In addition, in vitro susceptibility tests of the isolates to protective and systemic fungicides were performed. Fungicides were evaluated at three concentrations and mycelial growth reduction (MGR) and conidial germination inhibition (CGI) was estimated.
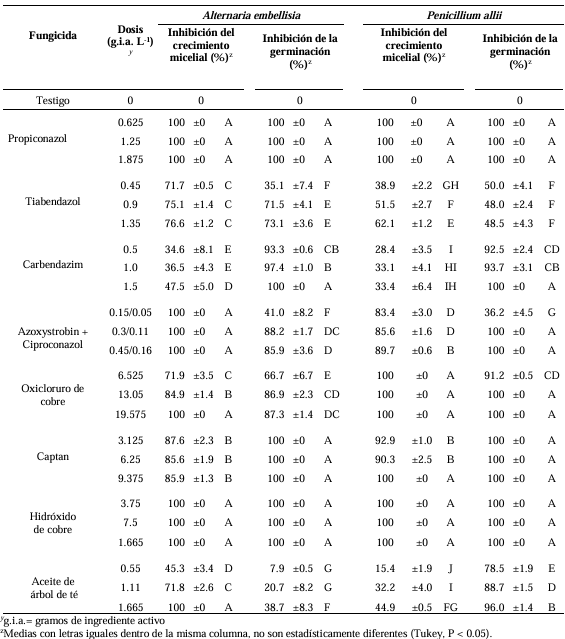
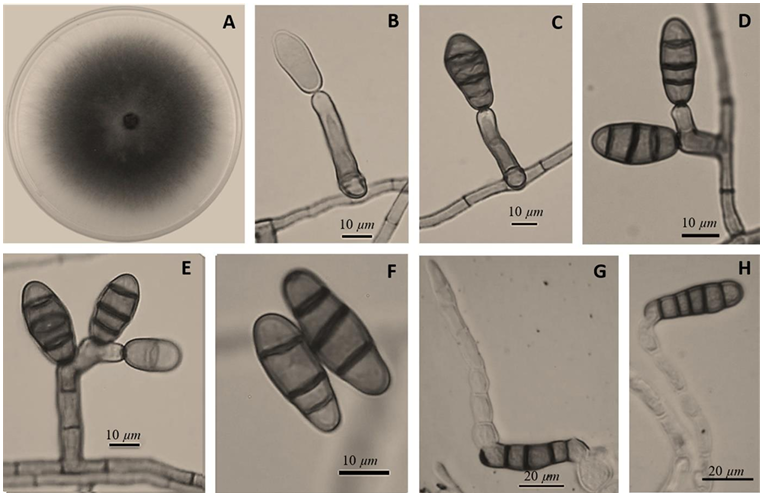
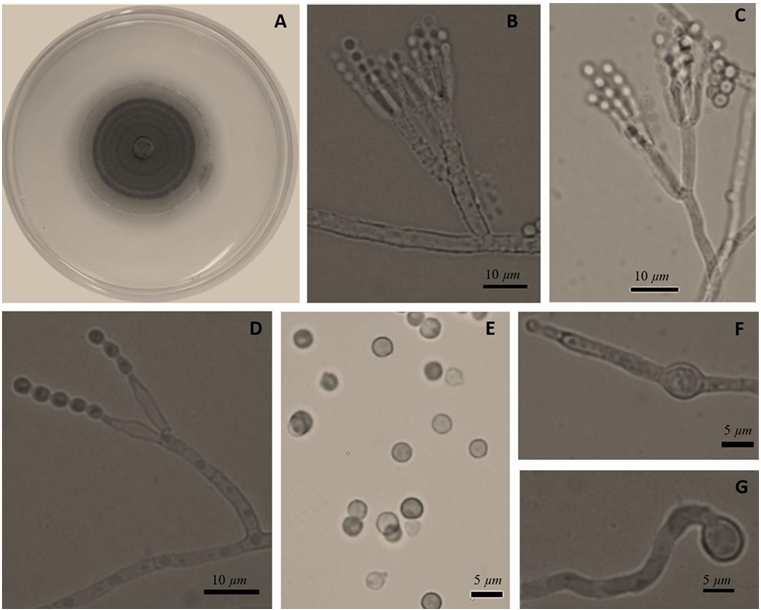
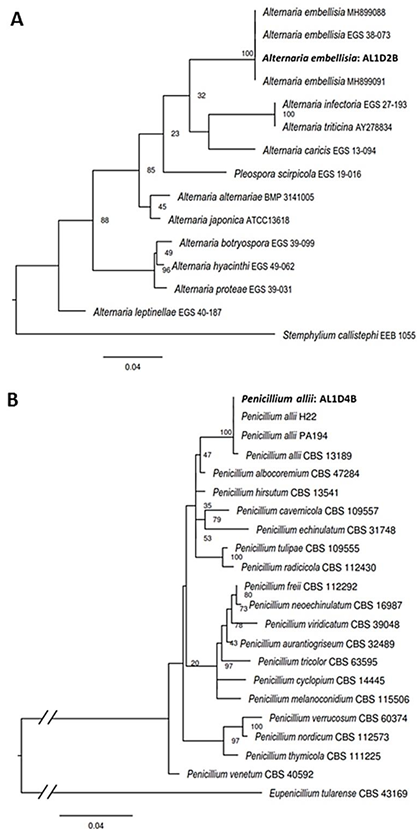
Results. The fungi Alternaria embellisia and Penicillium allii were identified in association with A. sativum bulbs with postharvest deterioration. P. allii showed the ability to develop internal infections from wounds; A. embellisia only showed growth on wounds. There were significant differences (p <0.0001) in the effectiveness of fungicides on the two species. Propiconazole and copper hydroxide inhibited 100% MGR and CGI in both fungi, at all doses evaluated.
Conclusion. P. allii is first reported as a causative agent of green garlic rot in Mexico. This study will serve as a basis for choosing control strategies and will contribute significantly to reducing economic losses in garlic production in this region.
Share this link via
Or copy link

byOscar Guillermo Rebolledo Prudencio, Wilberth Chan Cupul*, Guadalupe Moreno Zúñiga, Carlos Adrián Cruz Jiménez, Juan Carlos Sánchez Rangel
Received: 15/November/2024 – Published: 21/February/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-09
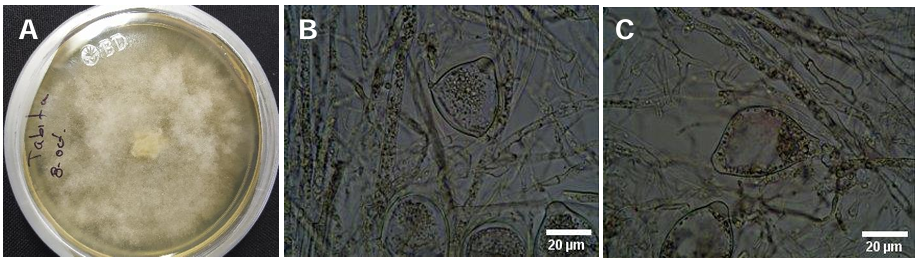
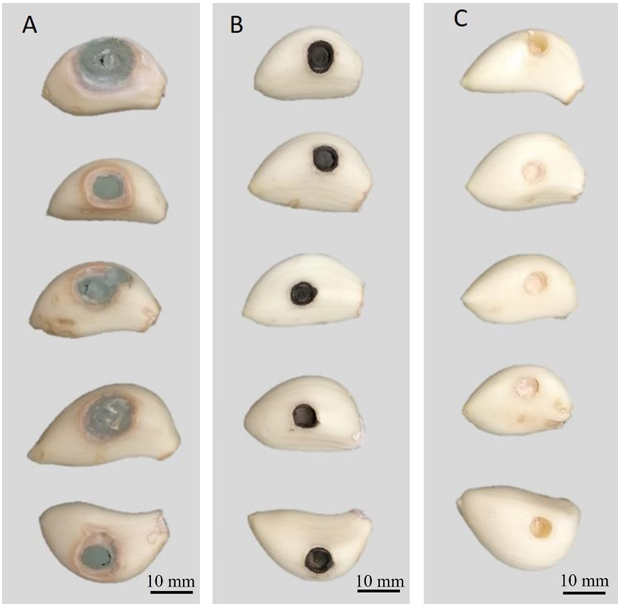
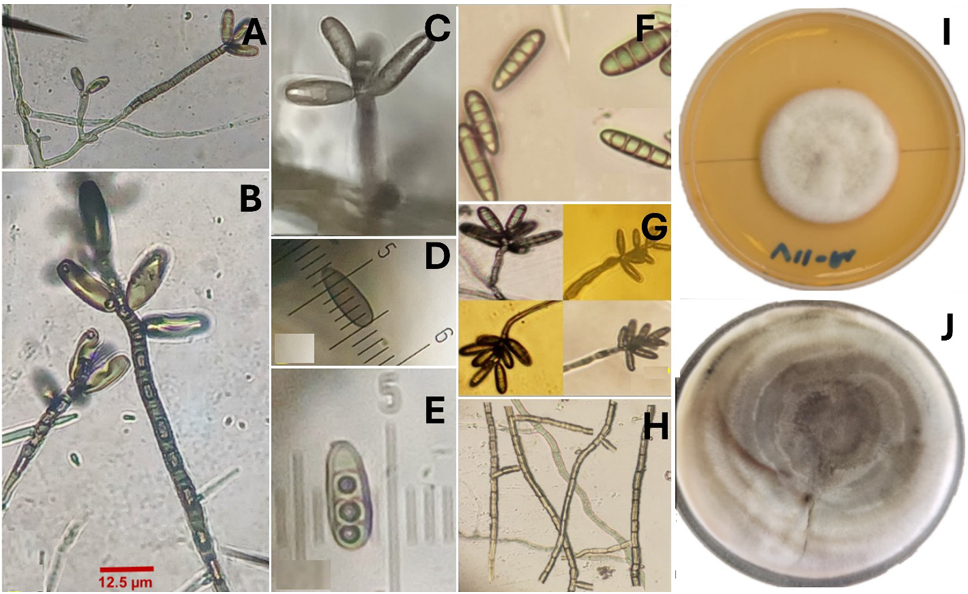
Abstract Background/objective. In Tecoman, Colima, Mexico, a leaf spot (LS) disease was detected with an incidence of 92.0% in Cocos nucifera hybrid Brazilian Green Dwarf (BGD). The objective was to characterize morphologically, molecularly and biochemically the fungus associated with LS in BGD coconut palm and evaluate its susceptibility to commercial biological fungicides.
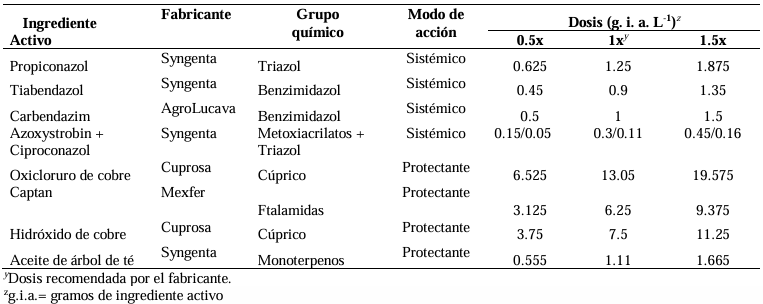
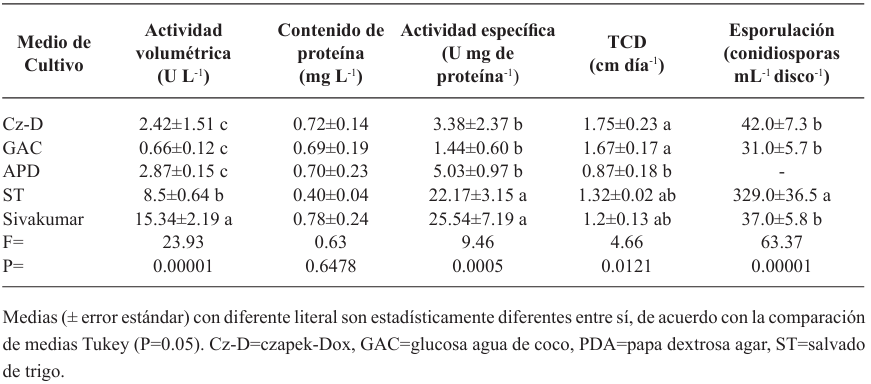
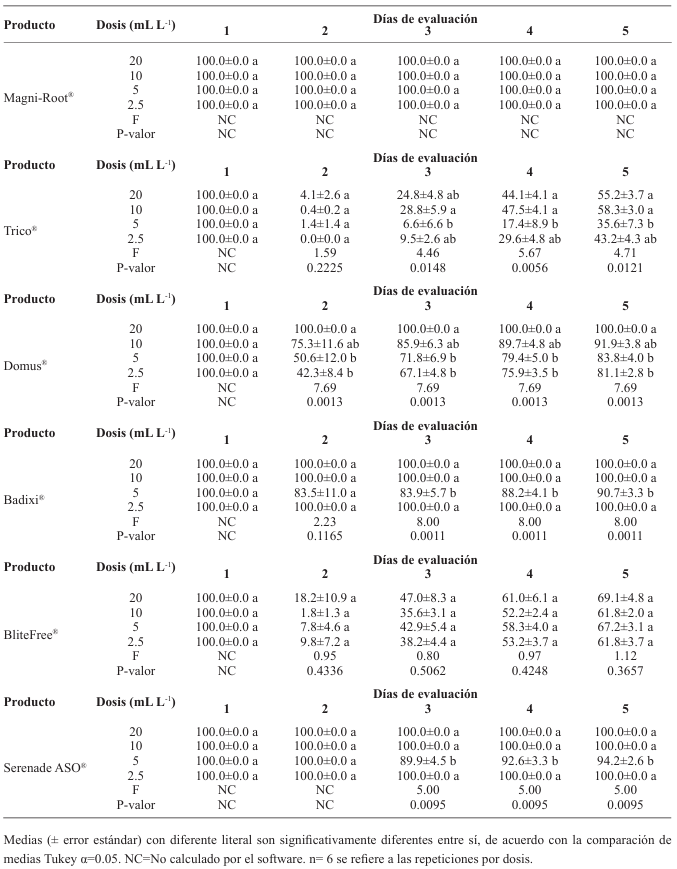
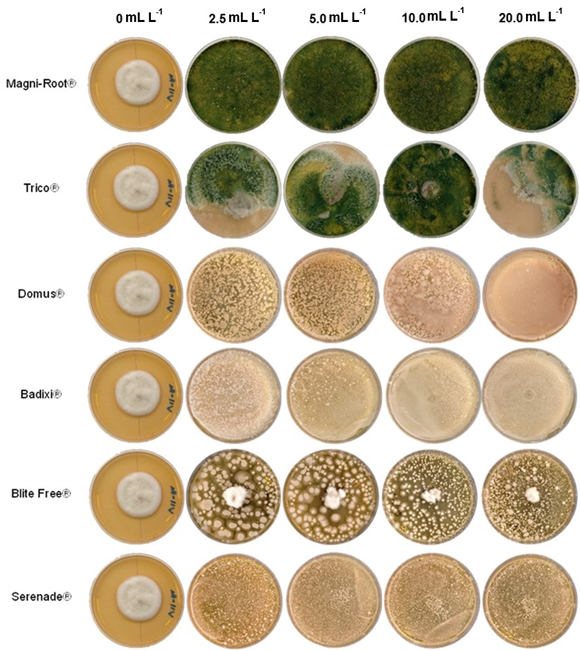
Materials and methods. The isolate was characterized morphologically and molecularly. Their growth, sporulation and laccase production were evaluated using different culture media. The in vitro mycelial inhibition and mean lethal doses (LD50) of commercial biological fungicides based on antagonistic fungi (Trichoderma harzianum and T. viride), bacteria (Bacillus subtilis and B. amyloliquefaciens) and actinobacteria (Streptomyces lydicus and S. jofer) were determined.
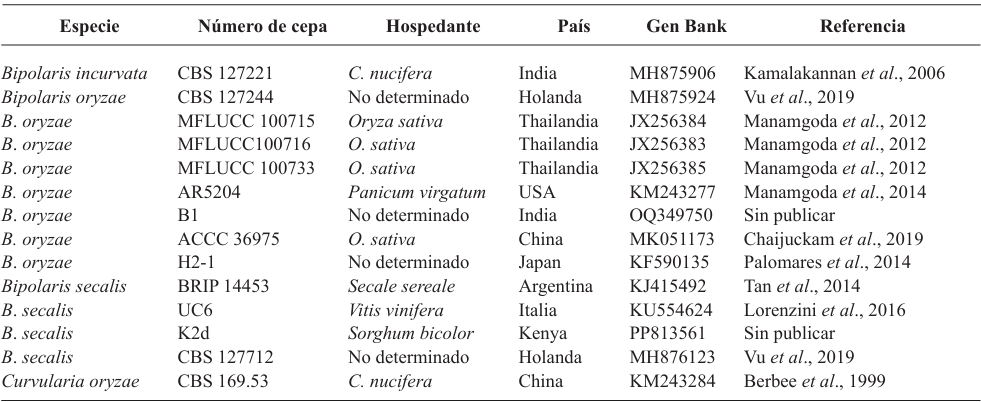
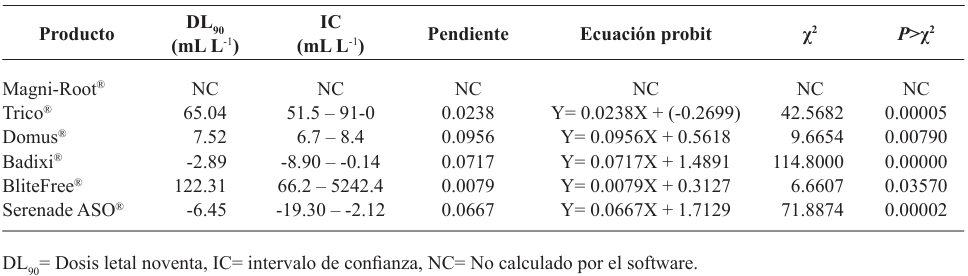
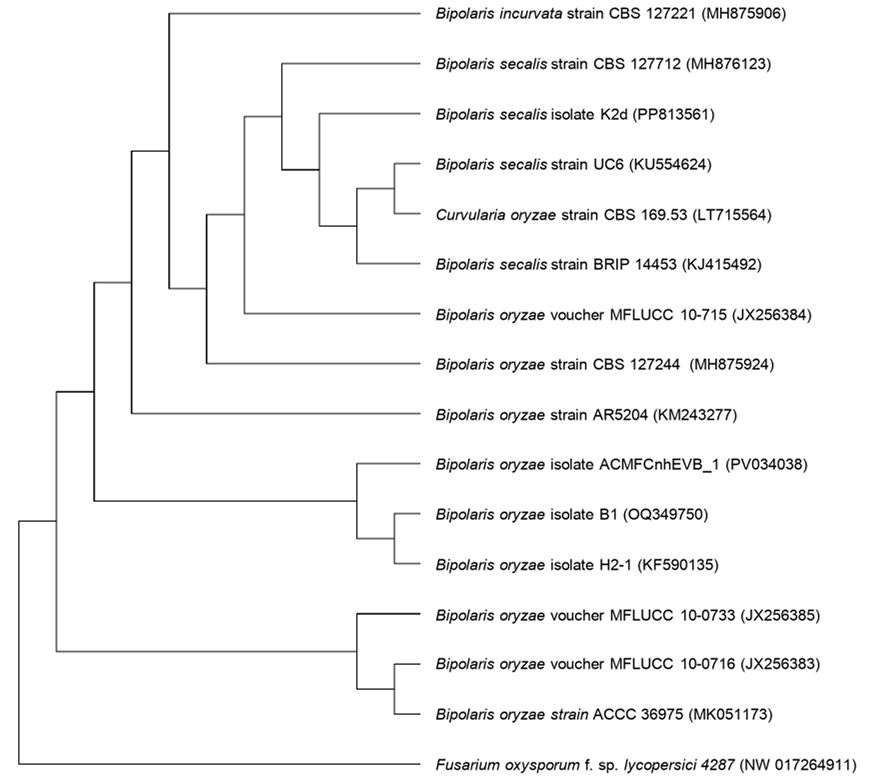
Results. Bipolaris oryzae was the associated agent of LS, it produces 25.54 and 22.17 U mg of protein-1 of laccase activity in the Sivakumar and wheat bran (WB) media. The WB medium allowed the greatest sporulation. Trichoderma harzianum inhibited B. oryzae at 100% in the four evaluated doses. B. subtilis and B. amyloliquefaciens inhibited B. oryzae at 100% at the highest tested doses (20 mL L-1).
Conclusion. Bipolaris oryzae is the associated agent of LS, it produced the highest laccase activity in Sivakumar and WB culture media. The highest sporulation and daily growth rate were in WB. T. harzianum stood out over T. viride by inhibiting B. oryzae growth by 100%. B. subtilis, S. lydicus and B. amyloliquefaciens were more effective against B. oryzae in vitro compared to S. jofer.
Share this link via
Or copy link

byMaría del Sol Cuellar Espejel, Evangelina Esmeralda Quiñones Aguilar, Gabriel Rincón Enríquez*, Rodolfo Hernández Gutiérrez, Juan Carlos Mateos Díaz, Sergio David Valerio Landa
Received: 15/November/2024 – Published: 13/February/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-25
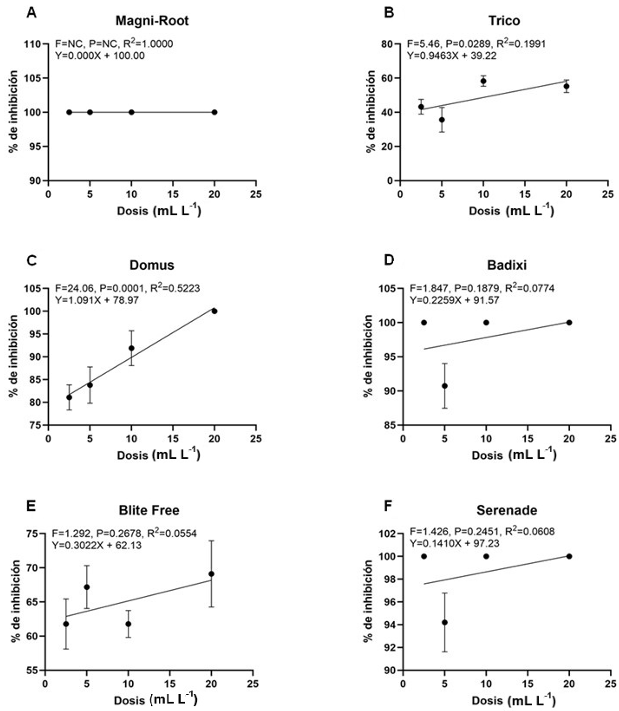
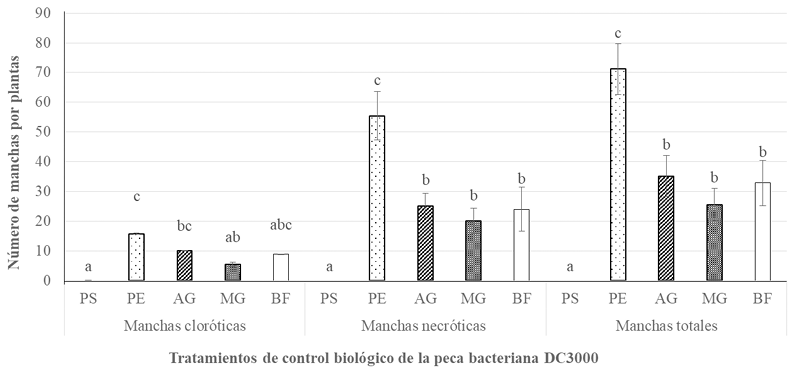
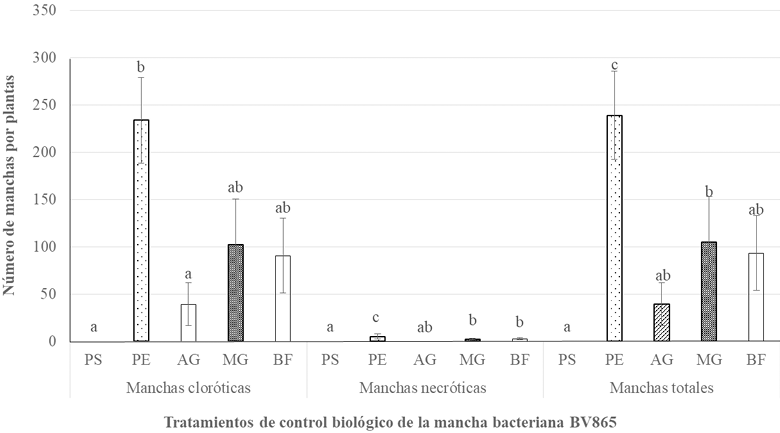
Abstract Background/Objective. Diseases such as bacterial spot in tomato (Solanum lycopersicum) and bacterial spot in chili pepper (Capsicum annuum) cause significant global economic losses. A sustainable alternative for their control is the use of protein inducers (Harpin proteins = Hrp) that activate plant defense responses by being recognized by the plant immune system, inducing defense mechanisms against pathogens. The objective of this research was to evaluate the biological effectiveness and optimal application dose of the biological inducer BioFensa (based on Hrp proteins), produced in a pilot plant, to control these diseases.
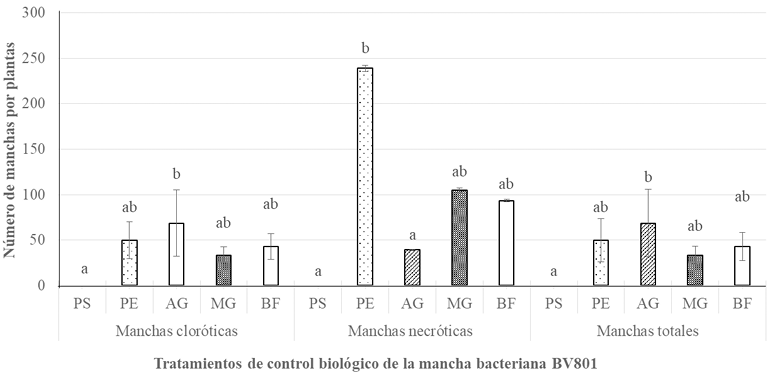
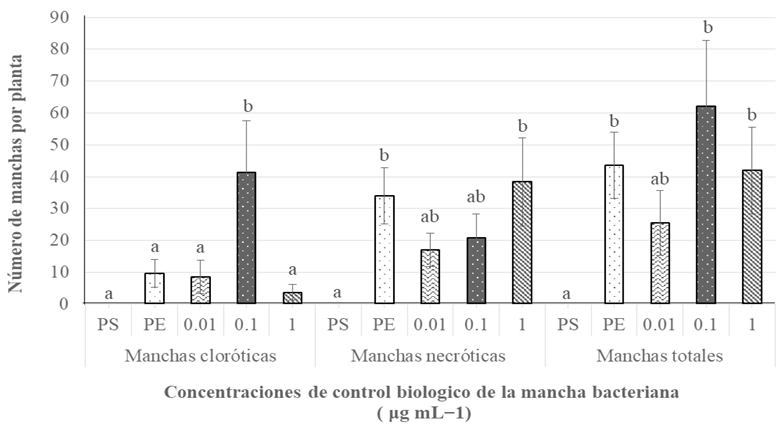
Materials and Methods. Three greenhouse experiments were conducted to evaluate the biological effectiveness of BioFensa (1 μg mL⁻¹). The protein inducer was tested for controlling bacterial spot (X. euvesicatoria strain BV865 [1] and BV801 [2]), as well as bacterial speck (P. syringae pv. tomato, strain DC3000 [3]). Each experiment included 5 treatments and 11 replicates. Additionally, an experiment was conducted to determine the optimal dose of BioFensa (0.01, 0.1, and 1.0 μg mL⁻¹) against X. euvesicatoria strain BV801, with 7 treatments and 8 replicates [4]. In the four experiments in total, plants were sprayed with BioFensa (3 mL per plant) 24 hours before infection, and symptoms were evaluated after 30 days by counting spots on the foliar tissue.
Results. BioFensa was effective in significantly reducing damage in chili and tomato plants (LSD, p≤0.05). At a high concentration (1 μg mL⁻¹), it prevented the appearance of spots on tomato plants by 53%, while for chili plants against strain BV865, it prevented spots by 60%. On the other hand, for chili plants against strain BV801, at low concentrations (0.01 and 0.1 μg mL⁻¹), symptoms were significantly reduced by 38-41%, whereas at a higher concentration (1 μg mL⁻¹), this effect was not maintained, suggesting a limit in the perception of inducers by the plants.
Conclusion. The results suggest that BioFensa has the potential to be an effective alternative to control diseases in horticultural crops such as tomatoes and chili peppers.
Share this link via
Or copy link
byWilberth Chan Cupul*, Osvaldo Villegas Guerrero, Juan C. Sánchez Rangel, Gilberto Manzo Sánchez, Marco T. Buenrostro Nava
Received: 15/November/2024 – Published: 13/February/2025 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-05
Abstract Background/Objective. Black Sigatoka (BS) is one of the main phytopathologies that reduces banana production in Mexico. Developing biological products based on antagonists is a predominant study activity. The production of conidiospores of Trichoderma spp. strains was evaluated in solid state fermentation using whole rice grains and cracked corn, and the effect of foliar applications of conidiospores against the BS epidemiology in banana cv. Great dwarf.
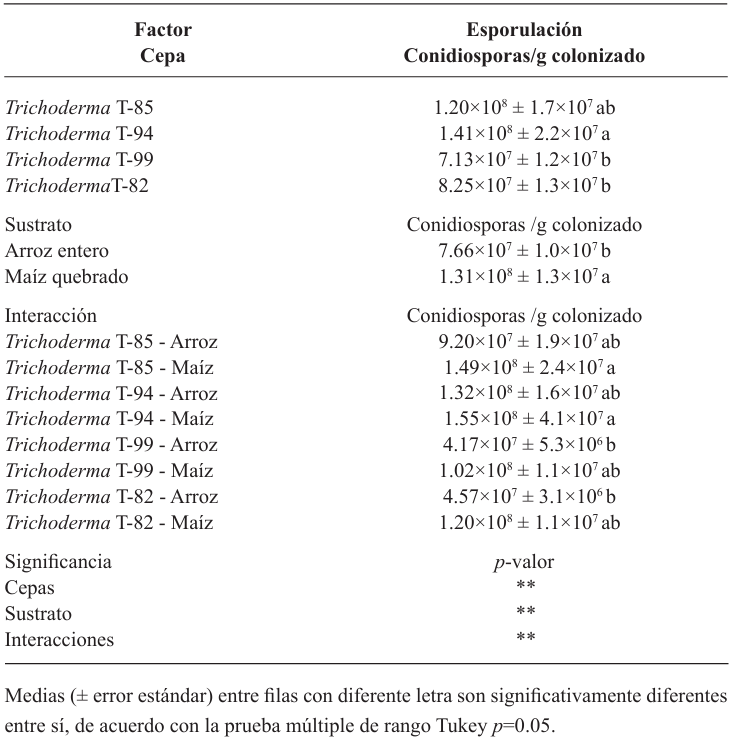
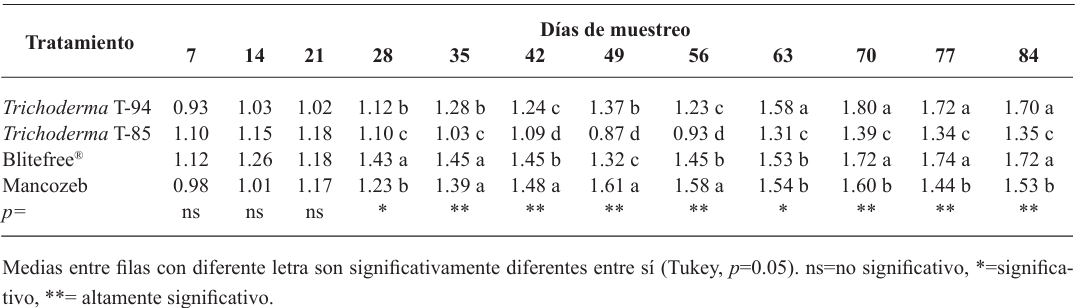
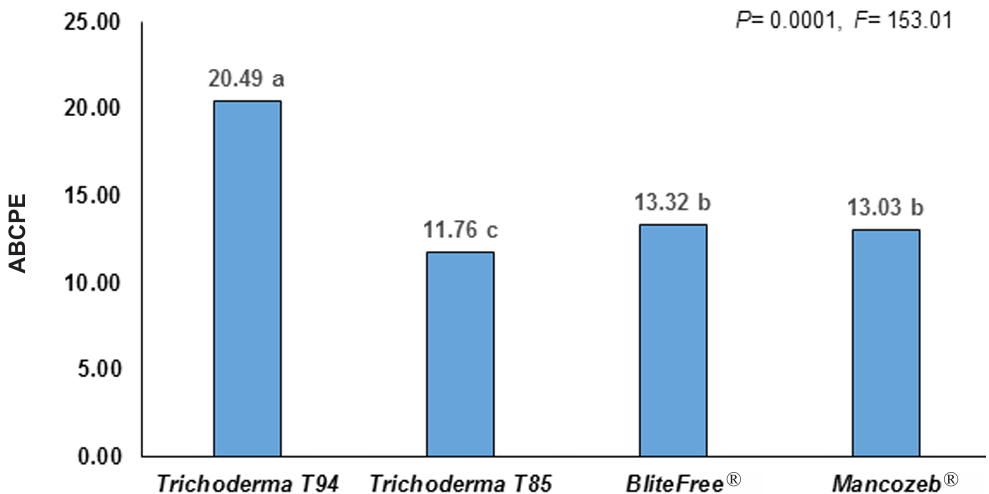
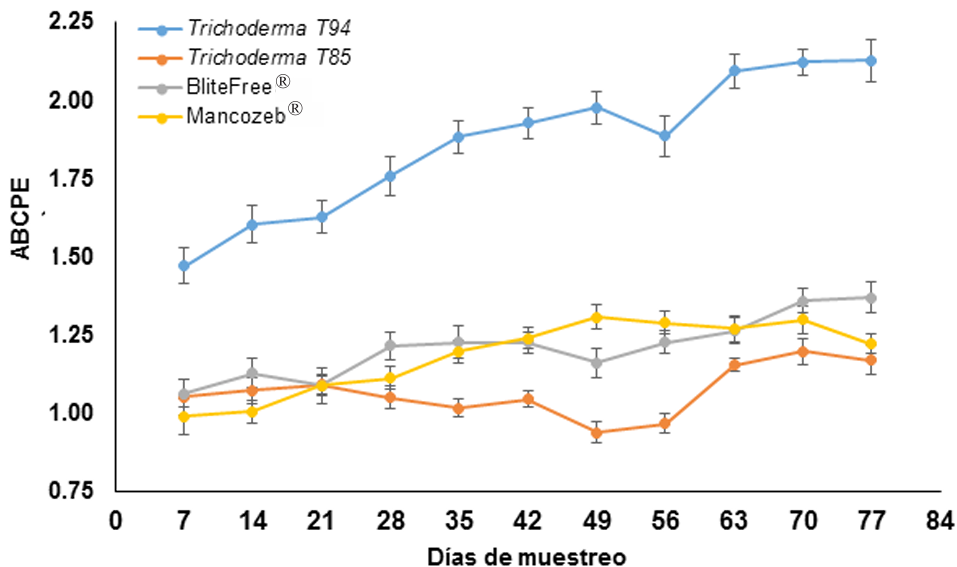
Materials and methods. In solid state fermentation, the yield of four strains of Trichoderma spp. (T-82, T-85, T-94 and T-99) in whole rice (WR) and cracked corn (CC) was evaluated, an A×B factorial design was used (A=strains and B=substrate). The two strains with the best yield (T-99 and T-85) and a Streptomyces spp. based product was applied in the field to evaluate the epidemiology of BS through the severity, weighted average of infection (WAI) and area under the disease progress curve (AUDPC), through a randomized block design.
Results. CC increased the yield of Trichoderma spp. in 71%, strains T-94 (1.41×108 conidiospores g-1) and T-85 (1.20×108 conidiospores g-1) achieved the highest yields. The T-85 strain reduced the severity, WAI and AUDPC of BS compared to applications of the chemical control “Mancozeb”.
Conclusion. CC was the best substrate to obtain greater yield in Trichoderma spp. T-94 and T-85. The weekly application of conidiosporas of Trichoderma T-85 reduces the severity, WAI and AUDPC of the SN in banana cv. Great dwarf.
Share this link via
Or copy link

byJeny Michua Cedillo, Gustavo Mora Aguilera*, Gerardo Acevedo Sánchez
Received: 30/July/2024 – Published: 31/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2406-7
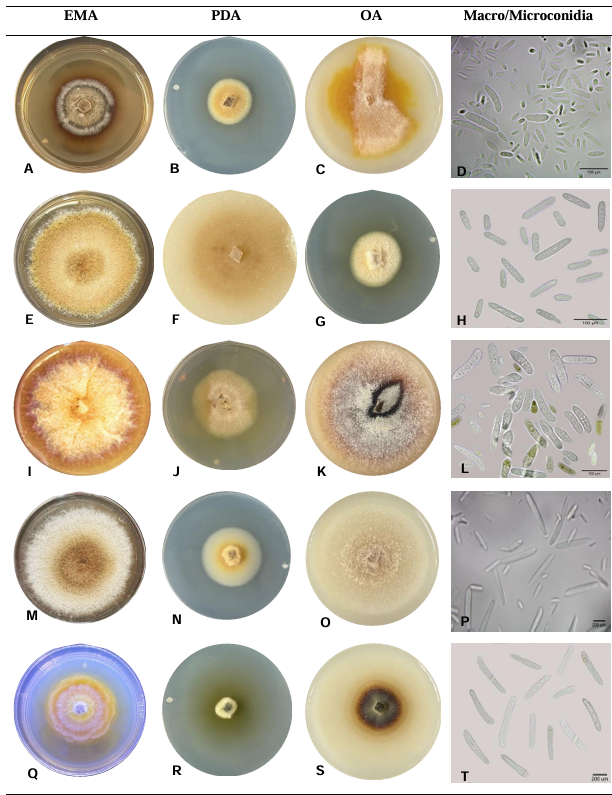
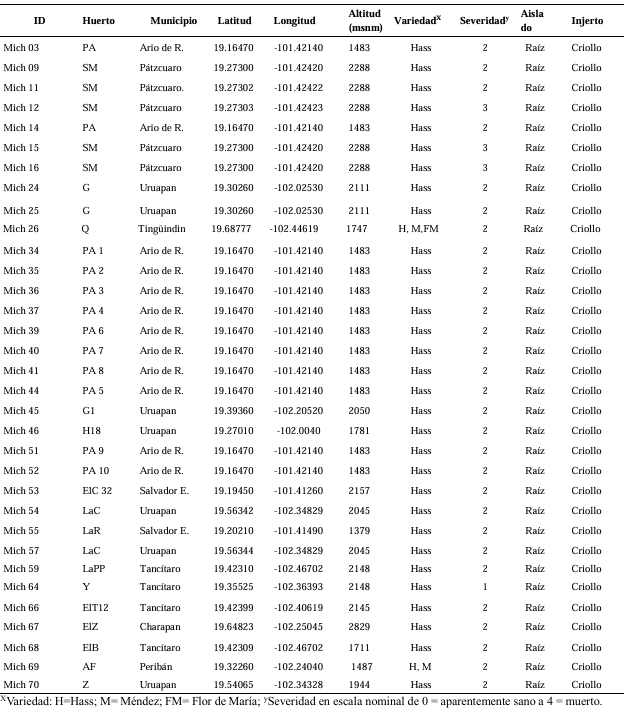
Abstract Background/Objective. Nectriaceae members increased regional occurrence in Michoacán since 2019. However, root species identity, geographic distribution, and association to other families are unknown. The objective was to characterize biological and molecularly species of Nectria associated with Persea americana.
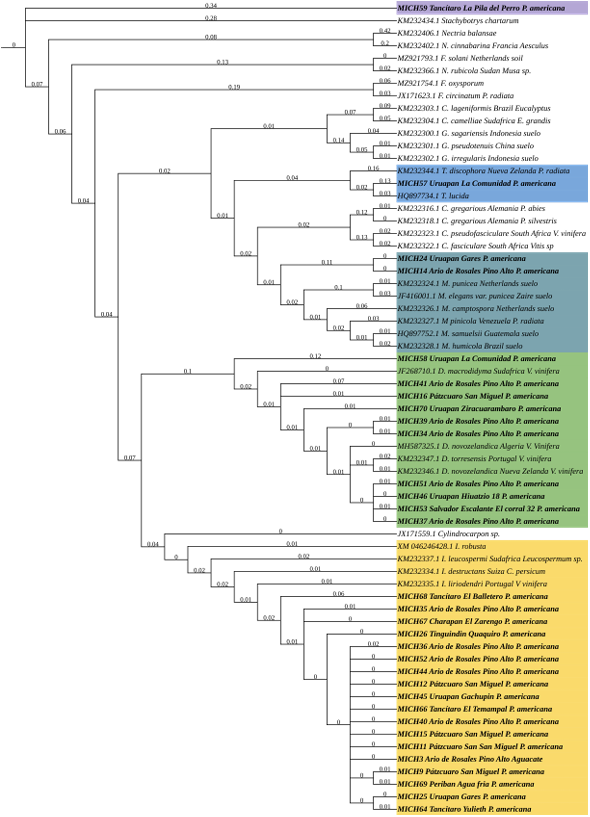
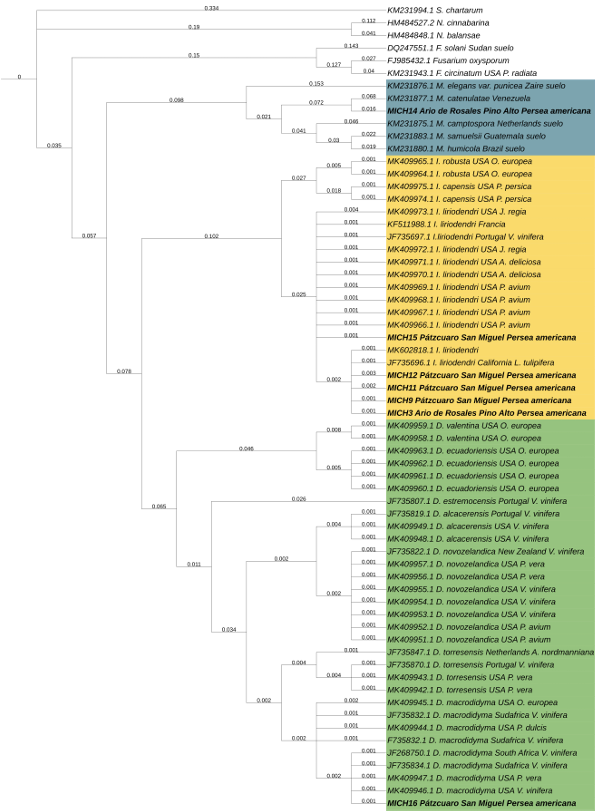
Materials and Methods. Seventy samples of wilt trees from 13 municipalities in Michoacán were processed. Thirty isolates selected based on epidemiological criteria were cultured in malt-agar, PDA, and oat-agar extracts to determine cultural and morphological characterization. Five morphotypes of Nectria with varying radial growth and brown coloration were obtained. From mycelial DNA, TEF 1-a and RPB2 genes were amplified, sequences were cleaned and aligned with SeqAssem and MAFFT, respectively. Bayesian inference and maximum parsimony phylogenetic algorithms were performed using PAUP 4.0 and MrBayes 3.2 complemented with 66 and 65 sequences from GenBank for TEF 1-a and RPB2, respectively. S. chartarum was used as the external species and four other Hypocreales.
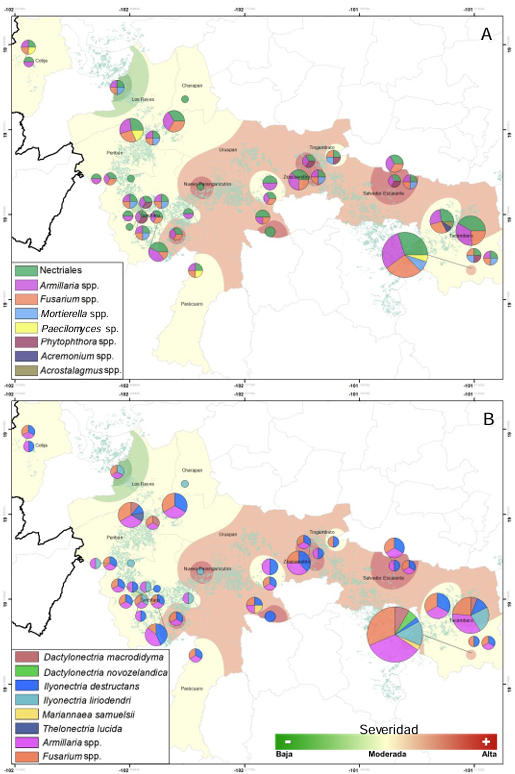
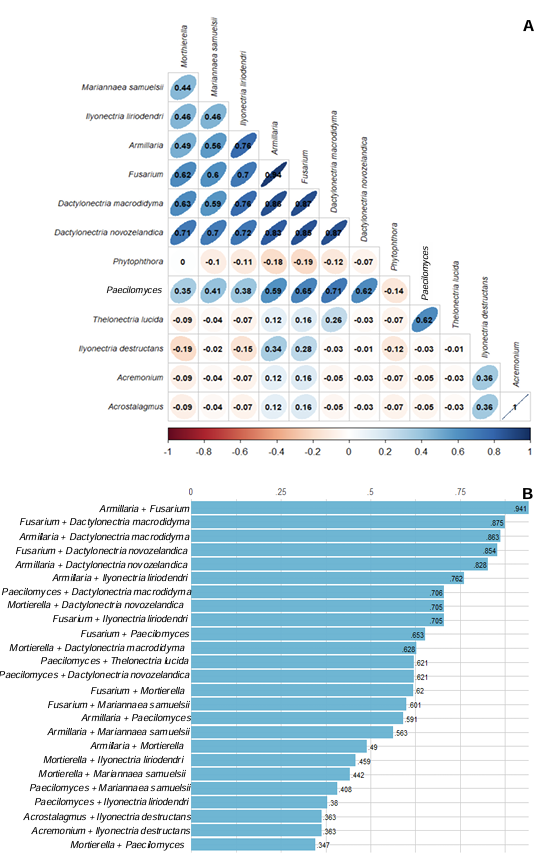
Results. Bayesian inference revealed greater phylogenetic consistency. Three genera and three species were identified with TEF 1-a (>94 % homology) and three genera and five species with RPB2 (>97 % homology) belonging to Ilyonectria (56 %), Dactylonectria (33 %), Mariannaea (6 %), and Thelonectria (3 %). Associations of Nectria were observed mainly to Armillaria (97.1 %), Fusarium (92.9 %), Paecilomyces (56.4 %), and Morthierella (47.3 %).
Conclusion. A decline syndrome in avocado trees associated with a fungal complex characterized by descending defoliation, wilt, reduced fruit size, and root necrosis is postulated. This is the first report of Nectria associated fungi in avocado trees in Mexico.
Share this link via
Or copy link


byGabriela Olivares Rodriguez, Juan Gabriel Angeles Núñez, Francisco Mondragón Rojas, Patricia Rivas Valencia, José Luis Zárate Castrejón, Luis Antonio Mariscal Amaro, Luis Febronio Díaz Espino, Talina Olivia Martínez Martínez*
Received: 10/July/2024 – Published: 31/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-28
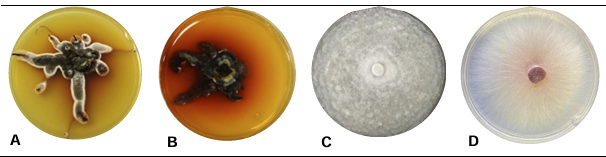
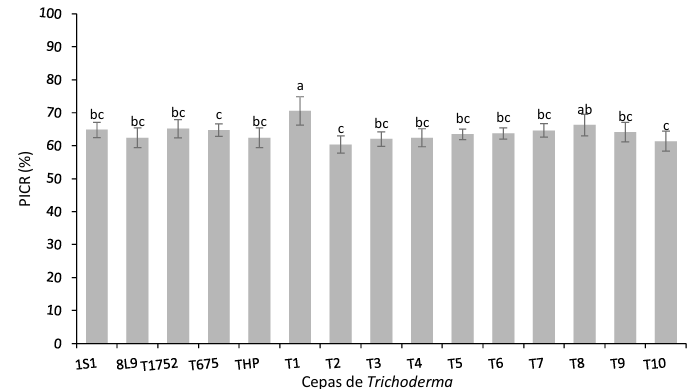
Abstract Background/Objective. The fungi Neopestalotiopsis sp. is an emerging pathogen that can cause losses of more than 70 % of production in strawberry crops. Due to this situation, is necessary to evaluate and implement control methods with low ecological impact. The objective of this work was evaluated the inhibitory growth of Neopestalotiopsis sp. using antagonist strains of Trichoderma sp. and fungicides used locally in the Bajio of Guanajuato, México.
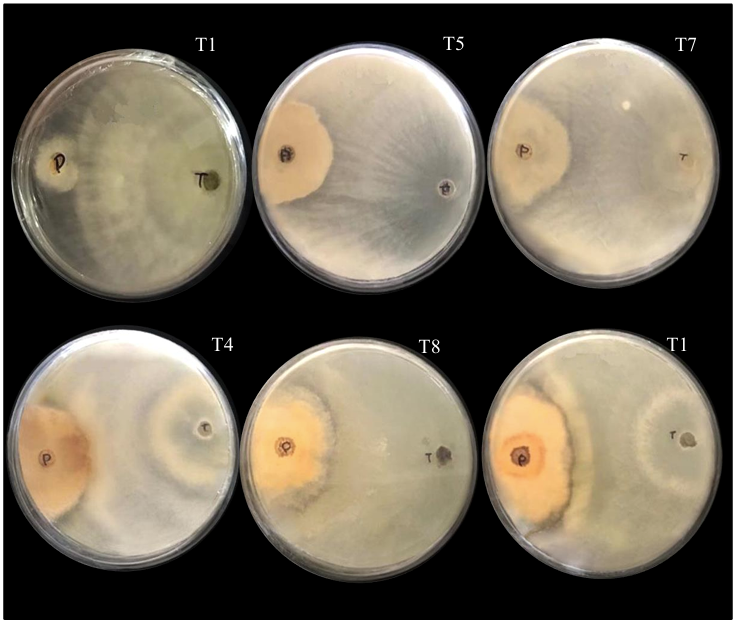
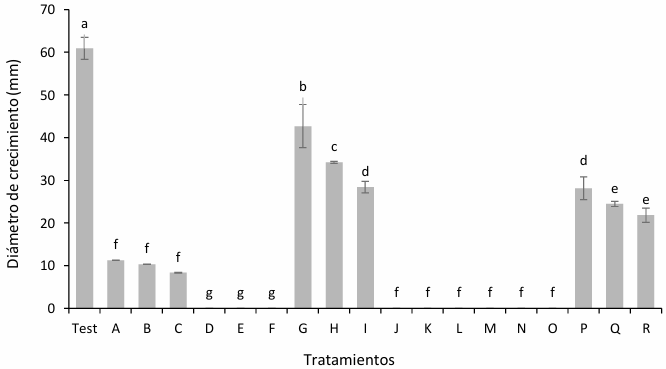
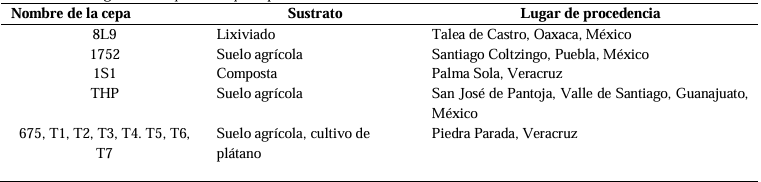
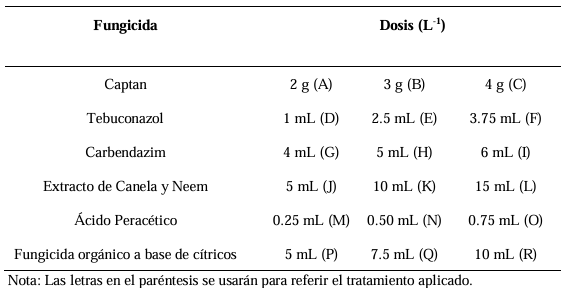
Materials and Methods. The pathogen was isolated in strawberry symptomatic plants. The morphological and pathogenicity characterization of the isolate was carried out. The Trichoderma strains were obtained from the biological collection of the National Forestry, Agricultural and Livestock Institute (INIFAP), Bajio Experimental Field (CEBAJ), and were confront in dual cultures with the pathogen, the percentage of radial growth inhibition (PRGI) was calculated at 120 h. In addition, five commercial fungicides were added to the growth medium and was calculated the growth diameter of the fungus.
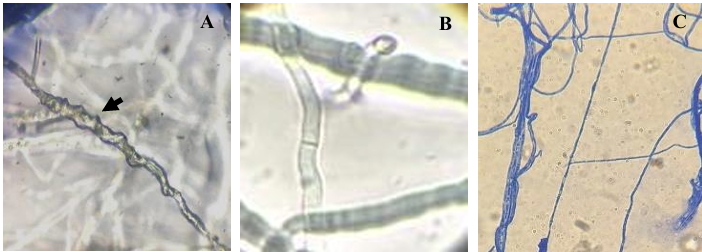
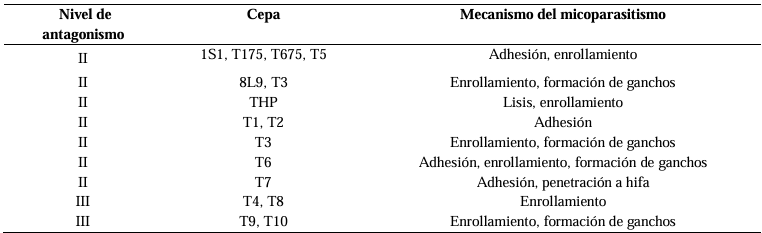
Results. The PRGI by Trichoderma were observed in a range of 63 to 70 %. The mechanisms of parasitism for Trichoderma were curling, adhesion and lysis to the pathogen hypha. The T1 strain was the greatest potential for controlling the pathogen, followed by T5 and T7. Three fungicides, Tecobenazole (100 mL 100 L-1), Cinnamon and Neem extract (500 mL 100 L-1), and Peracetic Acid (25 mL 100 L-1) had 100 % inhibited of fungal growth.
Conclusion. These results contribute to the knowledge on the control of Neopestalotiopsis sp. with the application of Trichoderma and the authorized products in Mexico
Share this link via
Or copy link



byAna Patricia Ibarra Valenzuela, Rosalba Troncoso Rojas, Alma Rosa Islas Rubio, Elizabeth Peralta, Herlinda Soto Valdez*, Hayati Samsudin
Received: 09/April/2024 – Published: 31/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-4
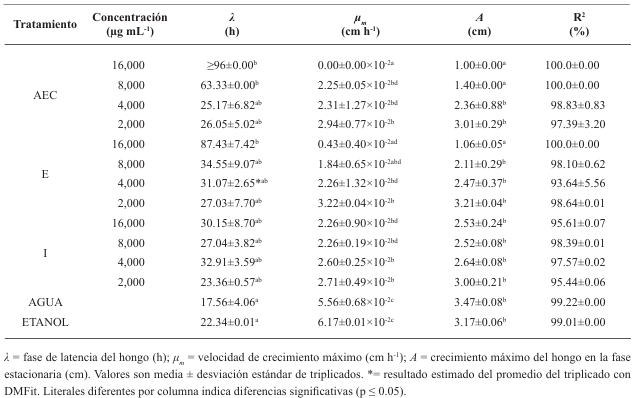
Abstract Objective/Background. Corn tortillas are a staple food in México that have a shelf life of 1-2 days at 25 °C due to fungal growth. A natural alternative for controlling fungal growth is clove essential oil (AEC) and its major components: eugenol (E), isoeugenol (I), and eugenyl acetate (AE). Objective: to evaluate the antifungal effect of AEC on the identified fungi present in corn tortillas.
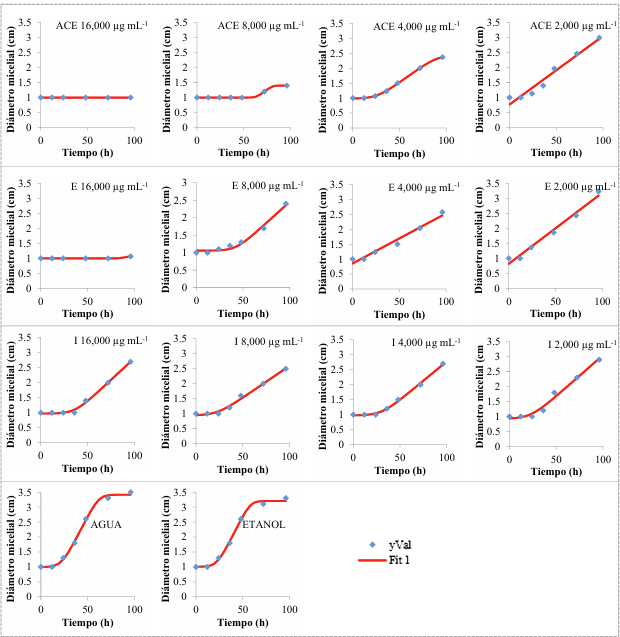
Materials and Methods. One kg samples of corn tortillas were obtained from the capitals of five states of Mexico (Sonora, Nuevo León, Michoacán, Oaxaca y Yucatán). Fungi were identified by their morphology and molecular biology. Moreover, the minimum inhibitory concentration (MIC) against AEC was determined. The effect of E, I, and AE on Aspergillus niger was evaluated with the Gompertz model.
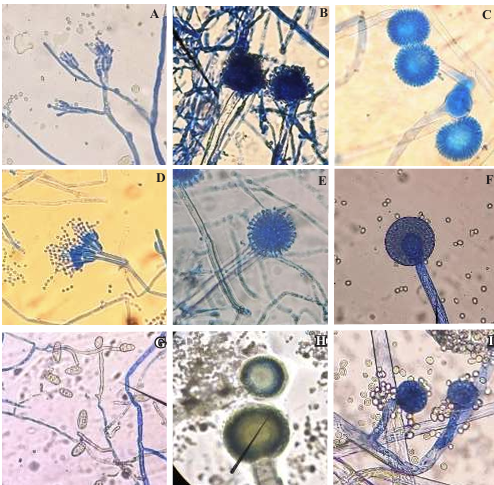
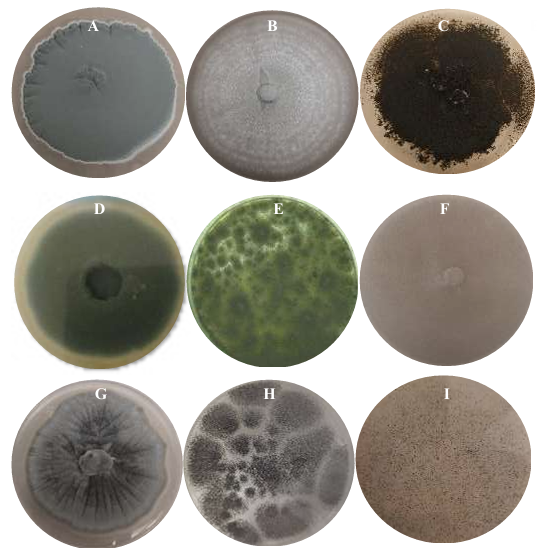
Results. Two fungi were isolated from corn tortillas purchased in Nuevo León, Sonora, Yucatán, and Michoacán, and one fungus from those purchased in Oaxaca. The following fungi were identified by molecular biology in corn tortillas: Aspergillus longivesica and Curvularia spicifera from Nuevo León; Aspergillus niger and Penicillium brevicompactum from Sonora; Aspergillus sp. from Oaxaca; Mucor sp. and Aspergillus flavus from Yucatán; Penicillium herquei and Curvularia racemosus from Michoacán. The MICs were 200, 400, 800, 400, 800, 400, 800, 800, and 400 µg mL-1, respectively. AEC, E, and I at a concentration of 800 µg mL-1 delayed the growth exponential phase of Aspergillus niger, while AE did not show any effect.
Conclusion. AEC could be a natural alternative for prolonging the corn tortillas′ shelf life.
Share this link via
Or copy link


byAugusto Gil Ceballos Ceballos, Yisa María Ochoa Fuentes*, Ernesto Cerna Chávez, Arely Cano García
Received: 18/April/2024 – Published: 31/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-5
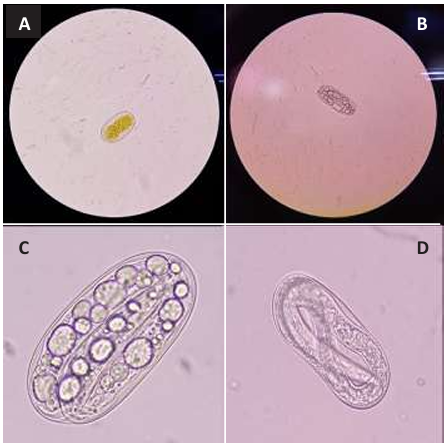
Abstract Background/Objective. Seed extracts from Canavalia ensiformis have shown both antiparasitic and repellent effects against pests. To evaluate the effectiveness of the extract combined with silicon dioxide nanoparticles (NPs) against Meloidogyne incognita eggs.
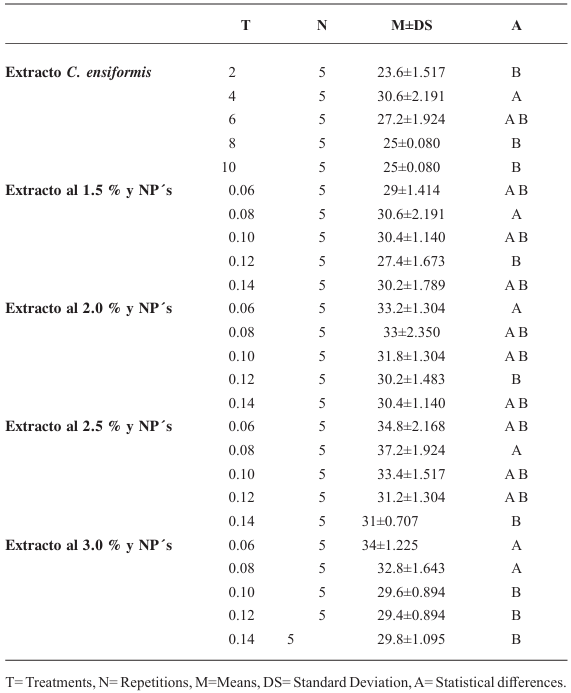
Materials and Methods. In vitro experiments were conducted to assess the effects of C. ensiformis seed extracts, alone and combined with silicon dioxide NPs, on M. incognita juveniles hatching. 150 eggs were used, and concentrations of 0, 2, 4, 6, 8, and 10 % of the extract were applied. Additionally, concentrations of the extract at 0, 1.5, 2.0, 2.5, and 3.0 %, each combined with NP concentrations at 0.06, 0.08, 0.10, 0.12, and 0.14 %, were evaluated.
Results. None of the treatments prevented more than 30 % of juveniles hatching. It was concluded that modifying the technique for obtaining C. ensiformis seed extract could have a complementary ovicidal effect; however, increasing the extract concentrations could serve as a medium for the proliferation of saprophytic fungi and other microorganisms.
Conclusion. The treatments did not show significant ovicidal effects.
Share this link via
Or copy link