
Share this link via
Or copy link


Article filters
Search Papers
byErika Janet Zamora Macorra, Norma Ávila Alistac*, Erika Lagunes Fortiz, Sergio de los Santos Villalobos
Received: 30/August/2023 – Published: 28/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-7
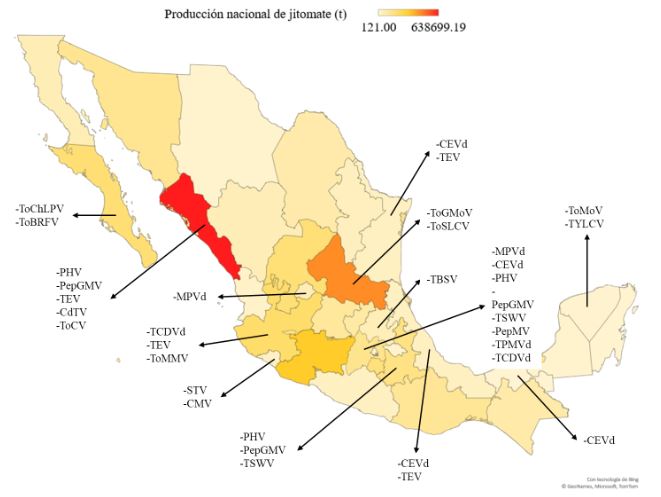
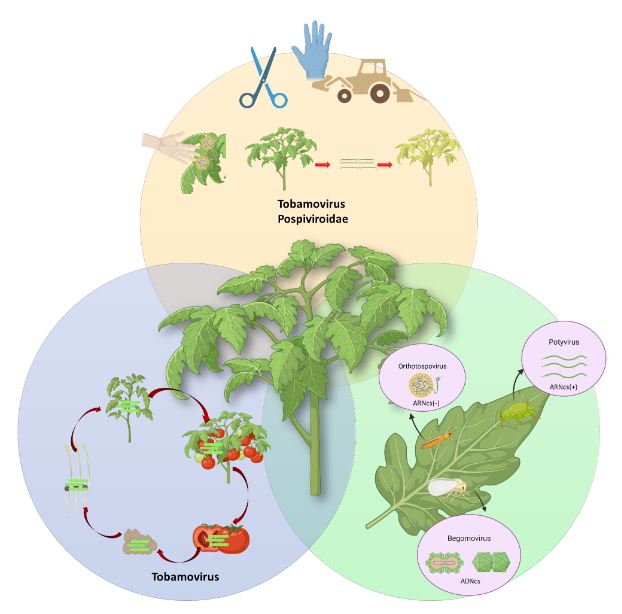
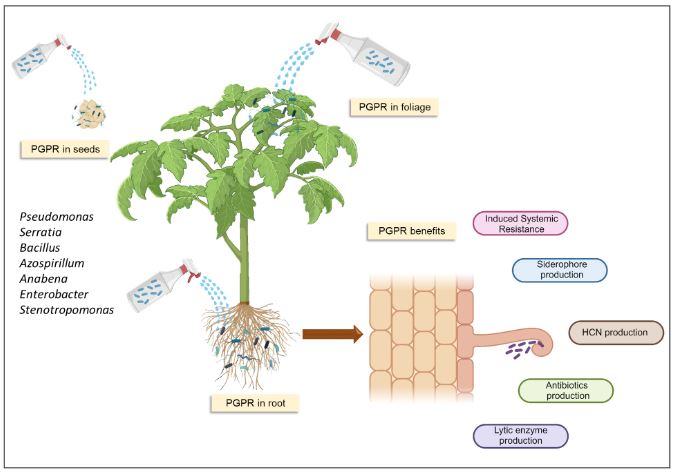
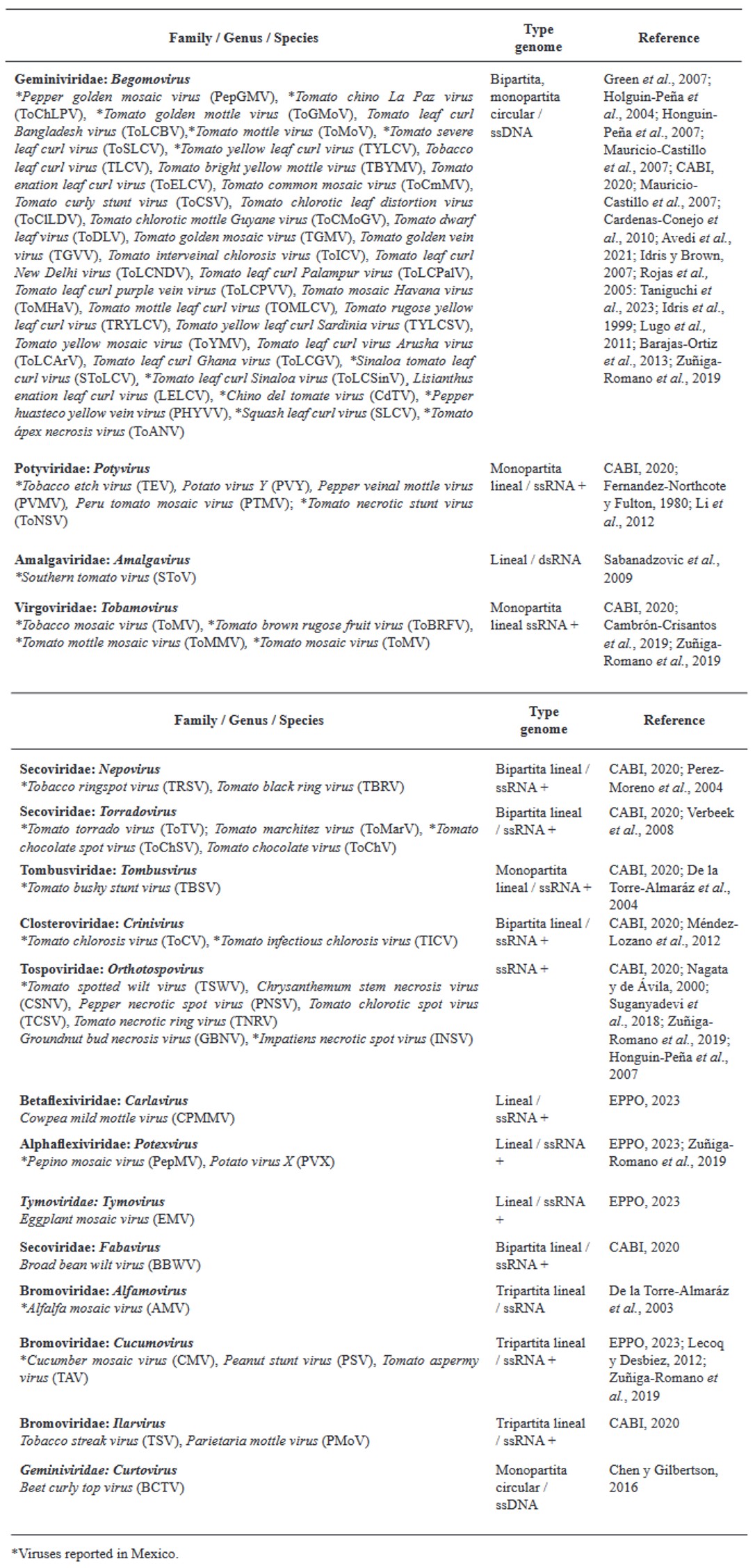
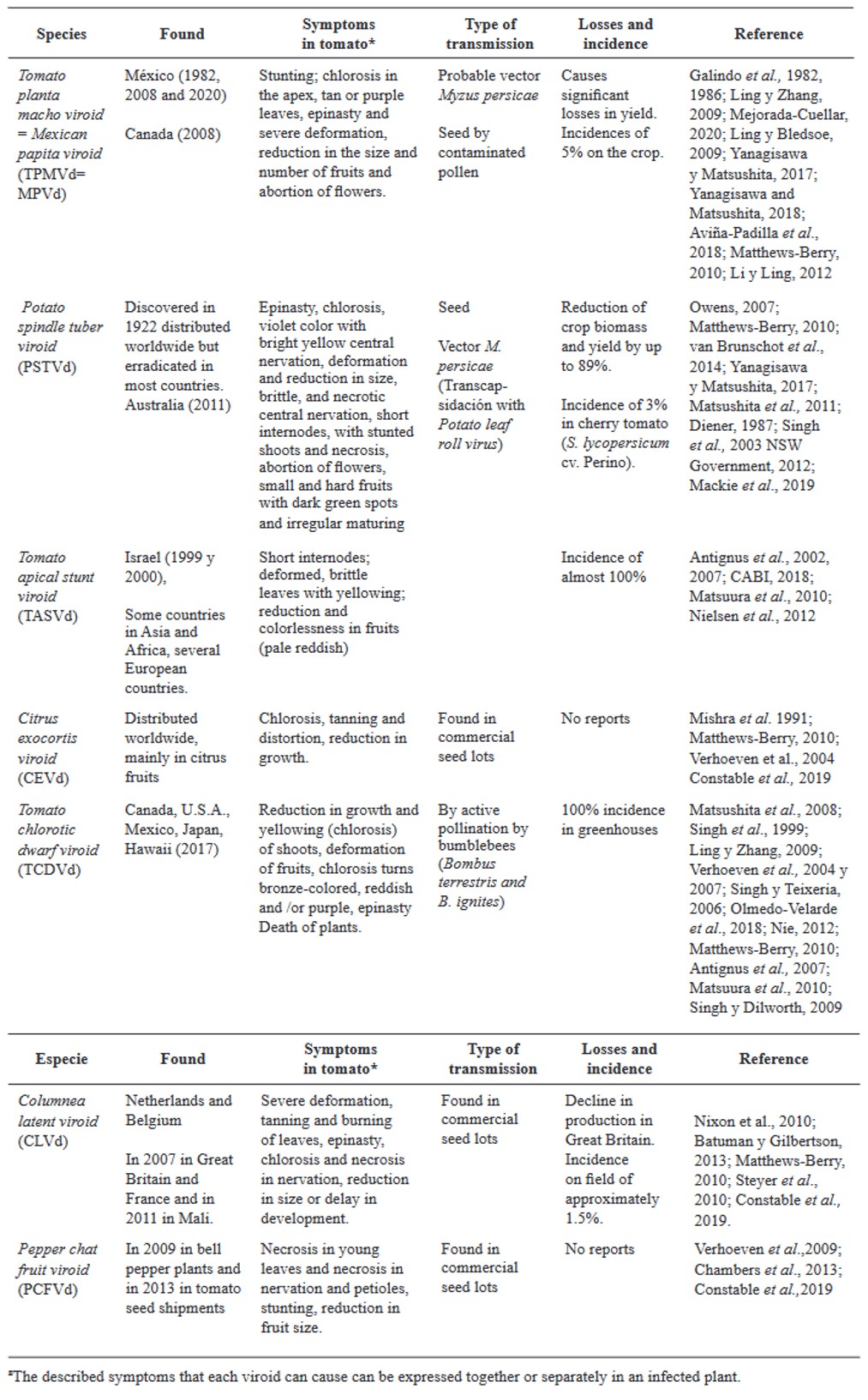
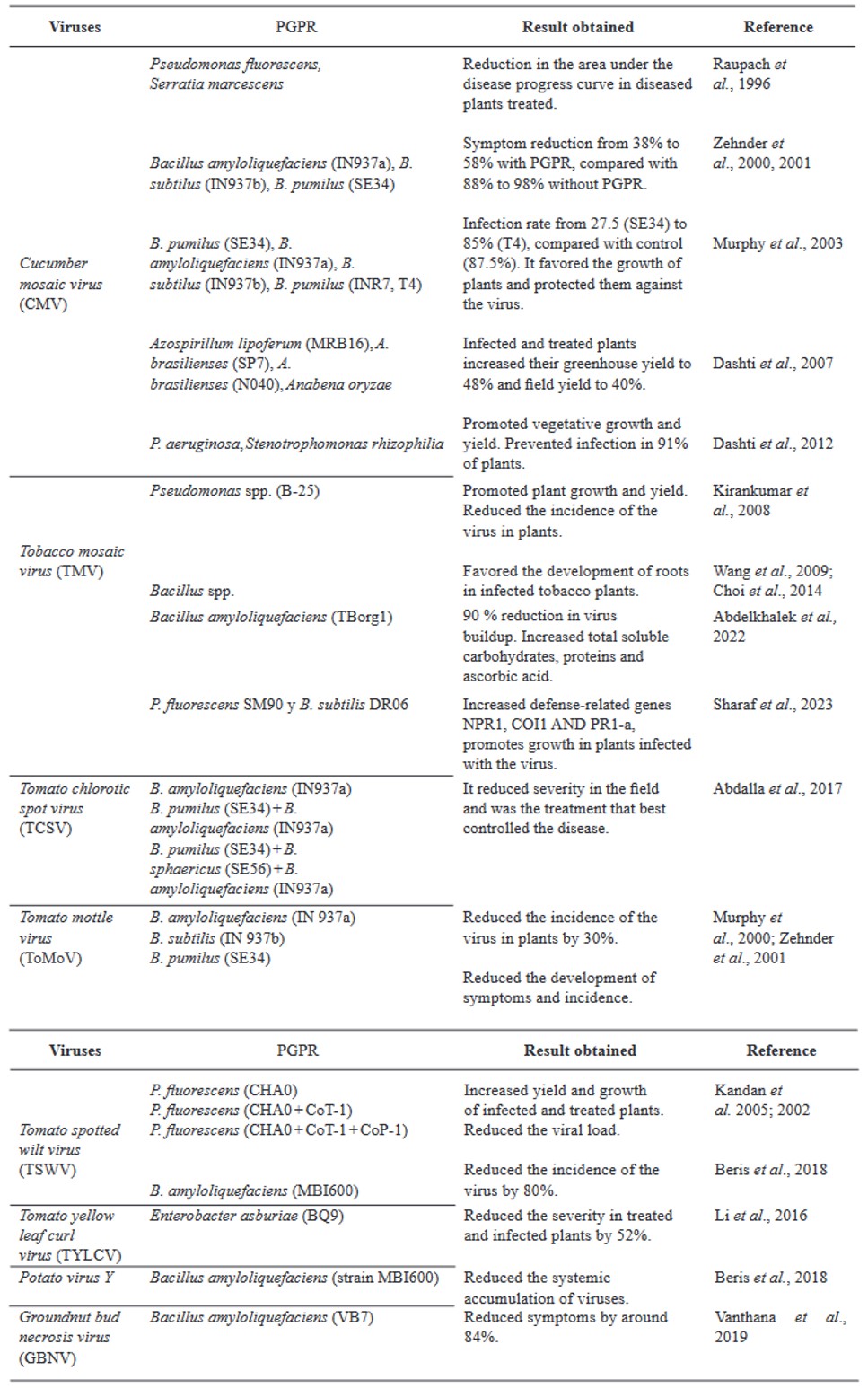
Abstract Viruses and viroids cause several diseases in tomato (Solanum lycopersicum) worldwide, generating important economic losses. About 312 viruses and seven viroids have been associated, of which more than 28 are present in Mexico. Therefore, the use of Plant Growth-Promoting Rhizobacteria (PGPR) can be an effective alternative for the management of viruses and viroids. The genera Pseudomonas, Bacillus, Azospirillum, Anabena and Stenotrophomonas have been implemented against main viruses reported in tomato: Cucumber mosaic virus (CMV), Tobacco mosaic virus (TMV), Tomato chlorotic spot virus (TCSV), Tomato mottle virus (ToMoV), Tomato spotted wilt virus (TSWV), Tomato yellow leaf curl virus (TYLCV), Potato virus Y (PVY), Groundnut bud necrosis virus (GBNV), with benefits in decreased incidence and severity up to 80 % and yield increase over 40 %. In Mexico, only Bacillus has been used. The use of PGPR is a strategy that could mitigate the impact of viral and viroid diseases and can be integrated into integrated management.
Share this link via
Or copy link

Weeds and ruderal plants as potential sources of inoculum for vegetable diseases in northern Sinaloa
byRubén Félix Gastélum, Gabriel Herrera Rodríguez, Karla Yeriana Leyva Madrigal, Guadalupe Arlene Mora Romero
Received: 31/July/2023 – Published: 28/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-4
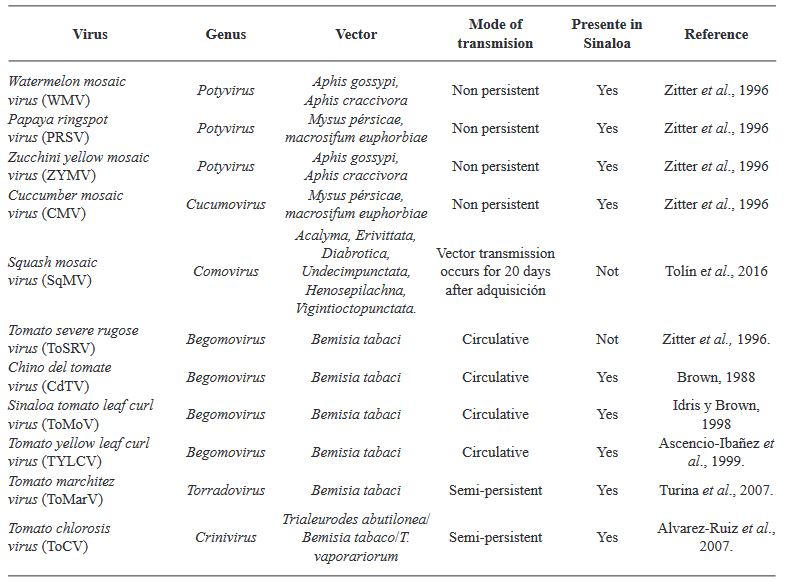
Abstract Weeds and ruderal plants of the families Cucurbitaceae and Solanaceae are addressed as potential sources of inoculum for the development of viral diseases such as Tomato apex necrosis virus (ToANV), zucchini (Zucchini yellow mosaic virus (ZYMV), Watermelon mosaic virus (WMV), Papaya ring spot virus (PRSV-W) and Cucumber mosaic virus (CMV). Reference is made to weeds and ruderal plants as potential sources of inoculum, including wild sunflower for powdery mildew (Golovinomyces spadiceus), wild tobacco for foliar blight (Alternaria spp.), black nightshade for leaf spot (Curvularia moehlemvekiae), Johnson grass for foliar blight (Alternaria sp.), and wild castor bean for foliar blight (Alternaria ricini) and wild melon for downy mildew (Pseudoperonospora cubensis). Future lines of multidisciplinary research focusing on the determination of pathogenicity in cultivated plants of viruses and fungi associated with wild plants and vice versa are proposed; the spatial-temporal distribution of wild plants that may serve as sources of inoculum, as well as the of potential insect vectors of viral diseases, should also be studied. The implementation of modern molecular techniques, such as High Throughput Sequencing, for the detection of phytopathogens is important. All this will contribute to the implementation of environmentally friendly strategies for disease control in agricultural crops in Sinaloa, for the benefit of the vegetable growers
Share this link via
Or copy link



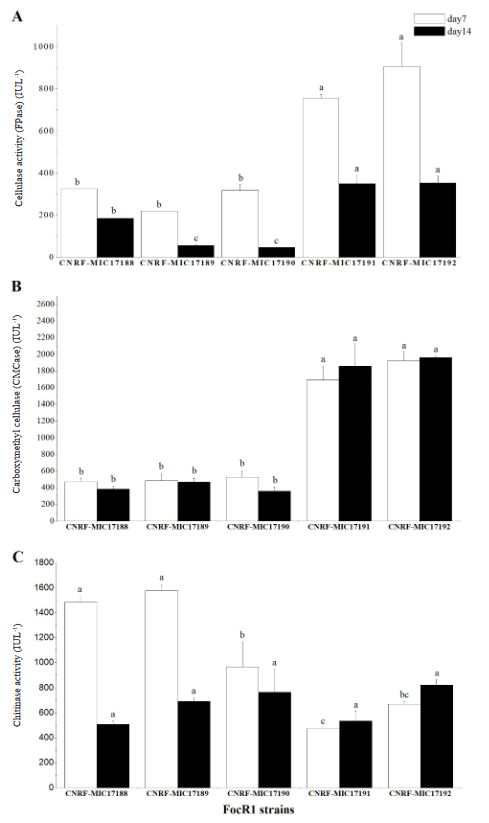
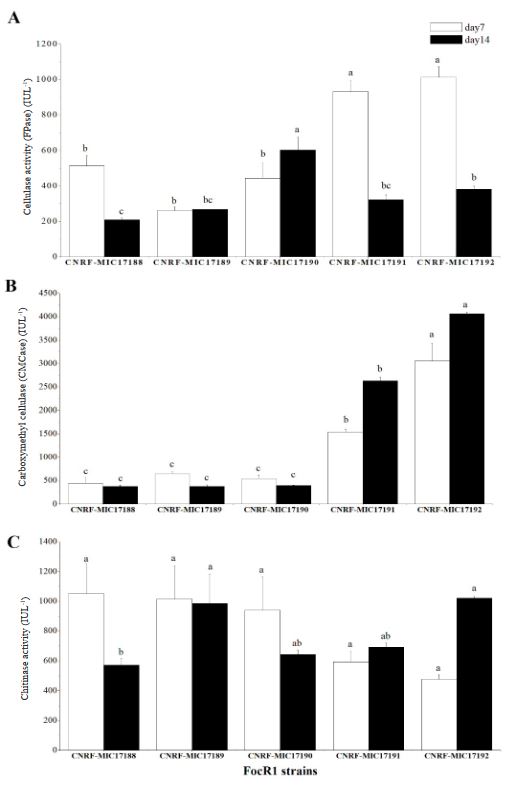
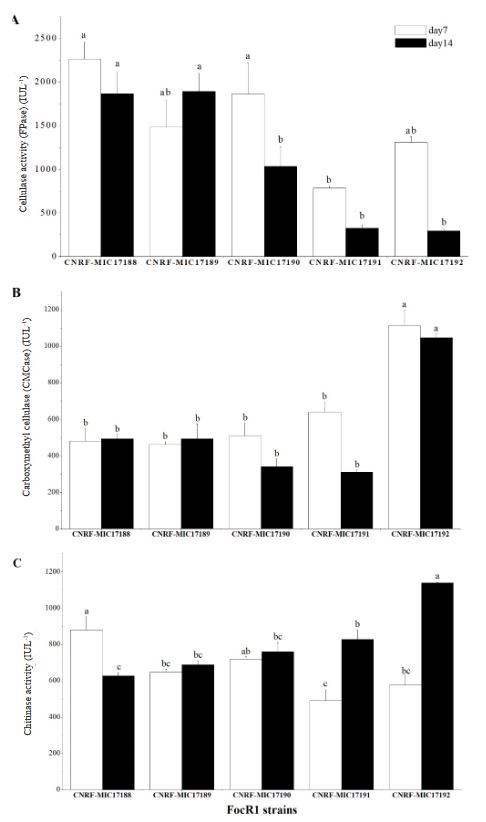
Cellulase and chitinase production by Fusarium oxysporum f.sp. cubense race 1 in submerged culture
byDulce Jazmín Hernández Melchor, Ronald Ferrera Cerrato, Clemente de Jesús García Ávila, Alejandro Alarcón*
Received: 09/July/2023 – Published: 29/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2307-2
Abstract Background/Objective. Fusarium has the capability to produce hydrolytic enzymes that can be used in the food and alcohol industries to break down natural organic compounds. This work studied the ability of Fusarium oxysporum f. sp. cubense race 1 (FocR1) to produce cellulases and chitinases enzymes in submerged culture using different carbon sources.
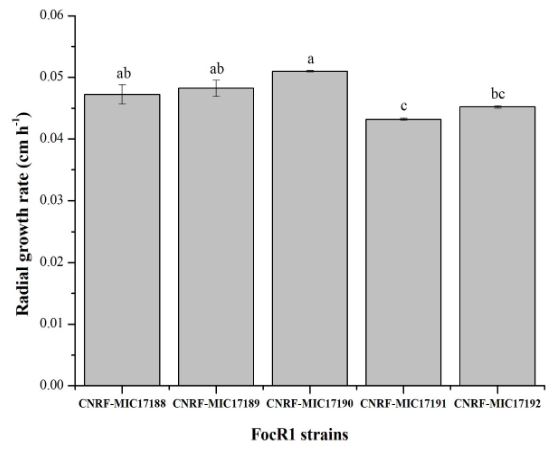
Materials and Methods. Five strains of FocR1 (CNRF-MIC17188, CNRF-MIC17189, CNRF-MIC17190, CNRF-MIC17191, and CNRF-MIC17192) were used in submerged culture for the degradation of three substrates [filter paper, newspaper, and chitin (Sigma®)], from where the radial growth rate (RGr) and the quantitative analysis of enzyme activities (FPase, CMCase and chitinase) were evaluated.
Results. The RGr of the five FocR1 strains oscillated in a range of 0.043 to 0.051 cm h-1. At 7 and 14 days, the five FocR1 strains produced cellulases and chitinases using the three substrates. Based on the statistical analysis, the strains CNRF-MIC17191 and CNRF-MIC17192 showed best results about enzymatic activities.
Conclusion. The five strains of FocR1 can be exploited as a commercial source of cellulases and chitinases, as well as potential candidates for bioconverting complex C-sources for further utilization in industrial processes
Share this link via
Or copy link
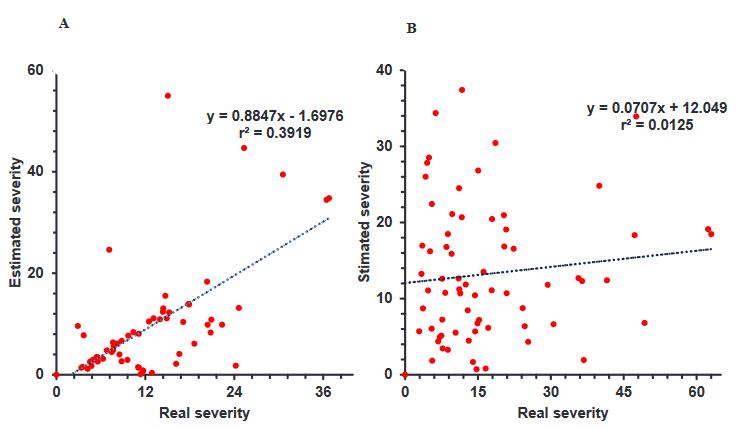
Diagrammatic scale to quantify the severity of Ascochyta blight in broad bean crops
byErnesto Alonso López Reyes, Álvaro Castañeda Vildózola*, Jesús Ricardo Sánchez Pale, Alejandra Contreras Rendón, Juyma Mayvé Fragoso Benhumea, Rómulo García Velasco
Received: 15/September/2022 – Published: 26/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2209-4
Abstract Background/Objective. The objective of this study was to design and validate a diagrammatic severity scale of brown spot on broad bean.
Materials and Methods. We collected 120 leaflets with different level of brown spot damage from commercial crops in the Toluca Valley, which were visually selected based on the expressed symptomology. Sixty leaflets were scanned for evaluation with the software APS PRESS ©Assess 2.0 to determine the real severity value for each leaflet.
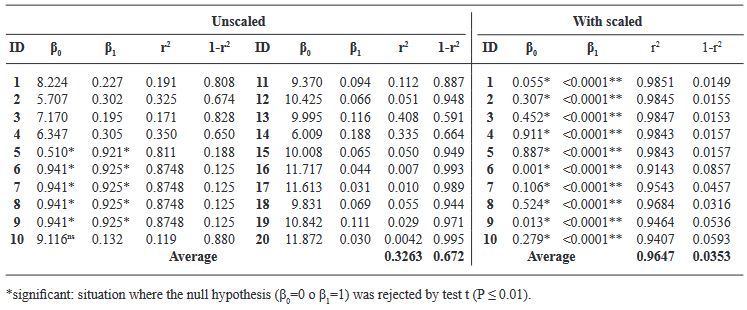
Results. The damage values allowed us to generate a diagrammatic scale consisting of six different classes: 0(0.0), 1(0.1-6.0), 2(6.1-10.0), 3(10.1-15.0), 4(15.1-40.0), 5(> 40.1-100). The leaflets were visually examined by evaluators who had no prior experience. The results from each evaluator were analyzed with a simple linear regression, obtaining r2 values from 0.0042 to 0.8748, β0 de 0.51 a 9.11, y β1 de 0.132 a 0.925. Using a scale, r2 values were obtained 0.9143 to 0.985, β0 de 0.001 a 0.911 y β1<0.001.
Conclusion. The generated diagrammatic severity scale was validated and reproducible, showing high reliability.
Share this link via
Or copy link

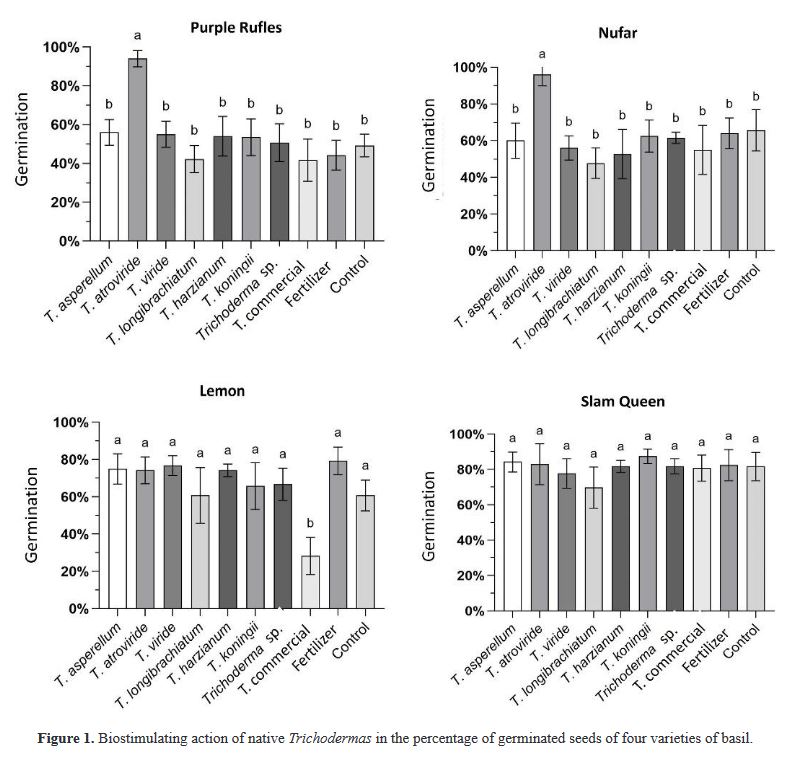
Biostimulant effect of native Trichoderma strains on the germination of four varieties of basil
byJuanita Guadalupe Hollman Aragón, Mirella Romero Bastidas*, Pablo Misael Arce Amezquita, Alejandro Palacios Espinosa
Received: 15/March/2023 – Published: 19/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2303-1
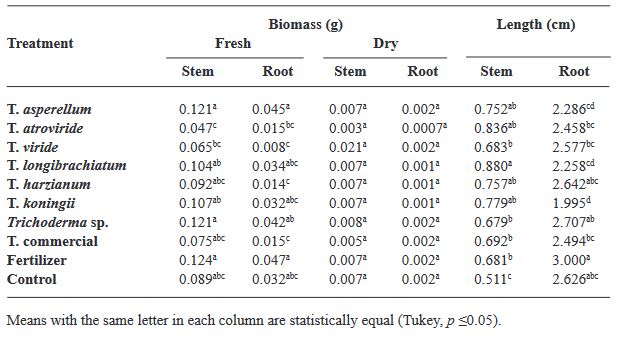
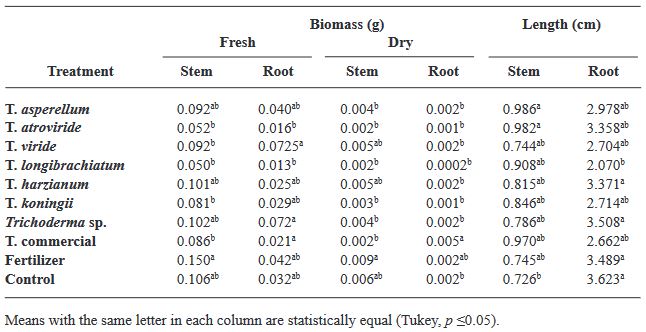
Abstract Objetive/antecedents. Trichoderma is an efficient tool as biostimulant in basil crop. However, only few species have been studied in specific cultivars. Therefore, the objective of this research was to evaluate the biostimulant efficacy of native Trichoderma strains on the germination and growth of four varieties of basil.
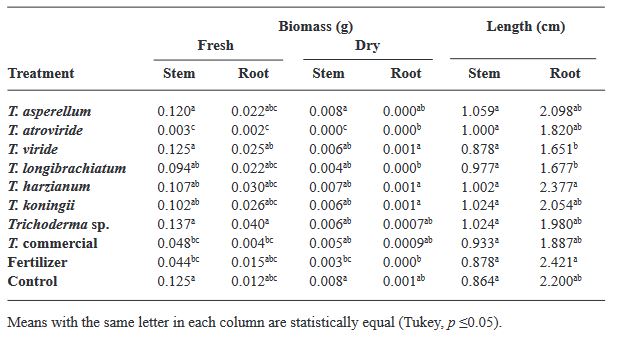
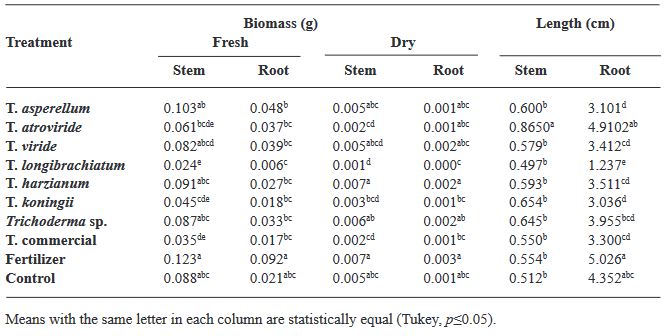
Materials and Methods. Seven strains of Trichoderma (T. asperellum, atroviride, viride, longibrachiatum, harzianum, koningii and Trichoderma sp.), a commercial Trichoderma (T. harzianum), synthetic fertilizer (T17) and the control were used in the study. 30 seeds of the Purple Ruffles, Lemon, Siam Queen and Nufar varieties were treated with a spore suspension of each Trichoderma. 48 h later, the seeds were sown and incubated at 28 °C with a 12 h light/dark photoperiod. The variables evaluated were; Rate and percentage of germination, biomass and length of seedlings.
Results. T. atroviride presented the greatest biostimulant effect on germination (95%). While T. asperellum registered an increased efficiency in biomass (≥ 0.120 g) and length (≥ 1.0 cm) of the plant in the four varieties. The action of commercial T. was lower in all cases.
Conclusion. This study demonstrated that the native strains of Trichoderma have a biostimulant effect on plants and are more effective than commercial species.
Share this link via
Or copy link

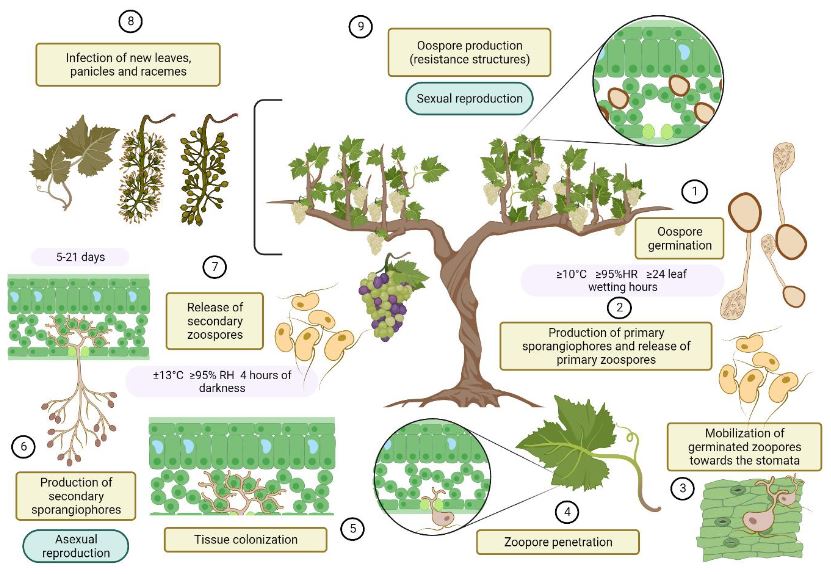
byDaniel Castrillo Sequeira, Rodrigo Jiménez Robles, Milagro Granados Montero*
Received: 18/September/2023 – Published: 27/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2309-3
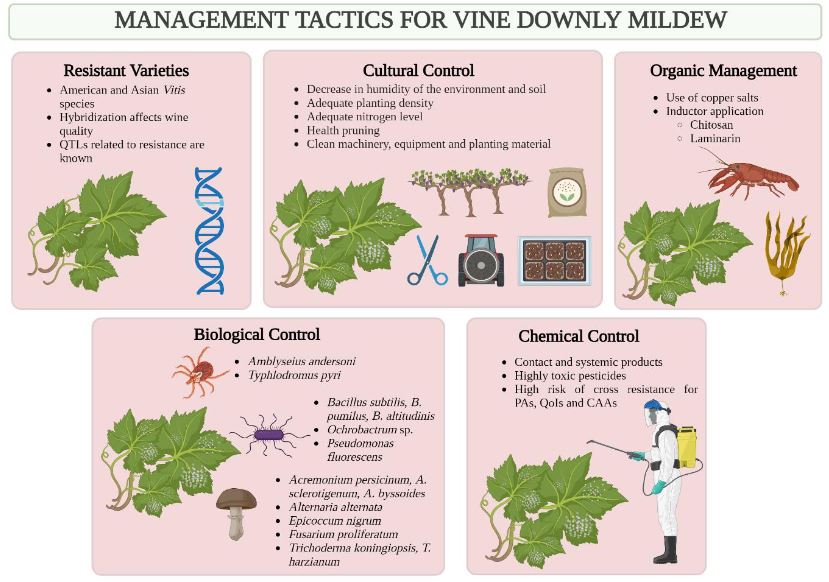
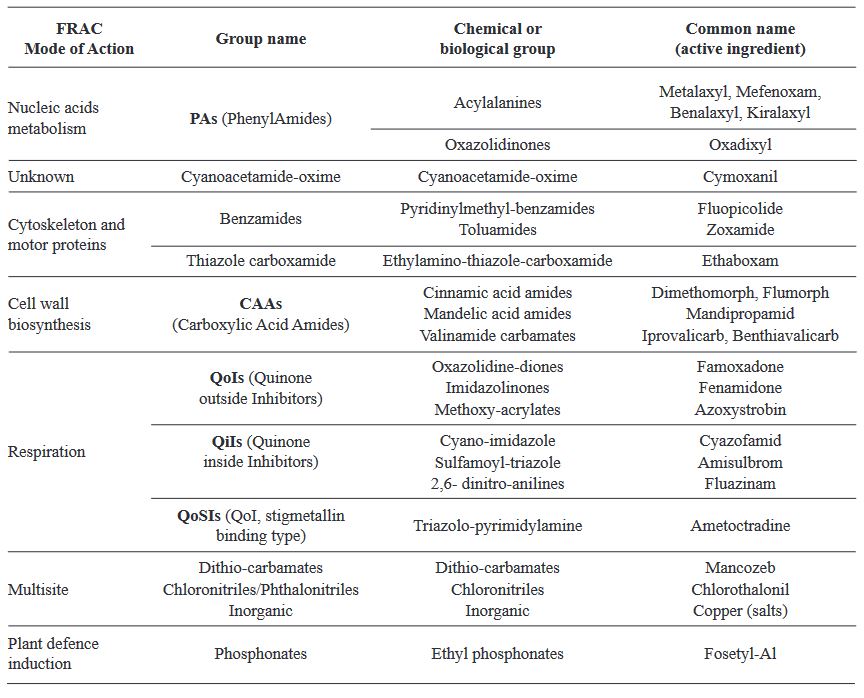
Abstract Viticulture is one of the oldest agricultural activities, and its exploitation has traditionally been limited to temperate climate zones, where the european grapevine (Vitis vinifera) and wine originate. Given the effects of climate change, more areas lose the capacity to grow this crop, and the tropics are presented as potential regions for this market. In Costa Rica, viticultural activity has been reported since the mid20th century, however, technical information on the crop is scarce. Downy mildew, caused by the oomycete Plasmopara viticola, represents one of the diseases with the greatest economic impact for viticulture worldwide, as well as the most limiting phytosanitary problem in Costa Rica. Under high humidity conditions, the development of the pathogen is accelerated, and the host remains susceptible throughout the crop cycle, which makes proper management of epidemics difficult. Chemical control is the most common management strategy around the world, however, the appearance of P. viticola populations with resistance to fungicides has been observed in most grape vine-growing areas, hence the search for more ecological alternatives is a necessity. Currently, Costa Rica does not have integrated management strategies that allow sustainable production, and there is only one registered product for protection against this pathogen. This situation justifies paying more attention to the investigation of this pathosystem
Share this link via
Or copy link

Induction of defense response mediated by inulin from dahlia tubers (Dahlia sp.) in Capsicum annuum
byJulio César López Velázquez, Soledad García Morales, Joaquín Alejandro Qui Zapata*, Zaira Yunuen García Carvajal, Diego Eloyr Navarro López, Rebeca García Varela
Received: 10/May/2023 – Published: 29/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2305-2
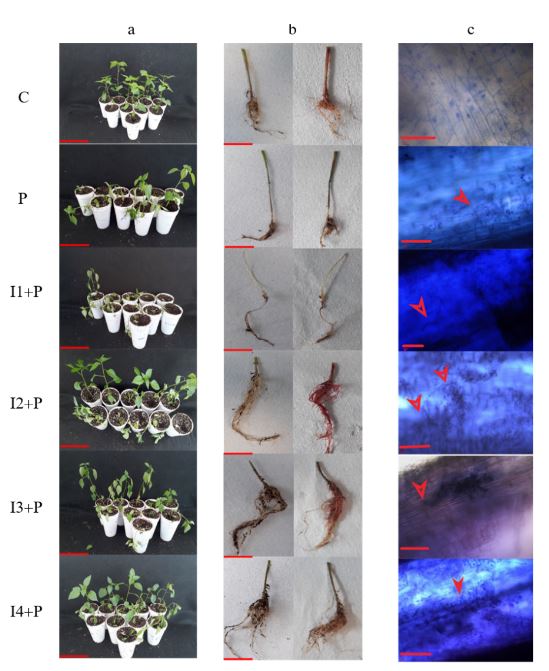
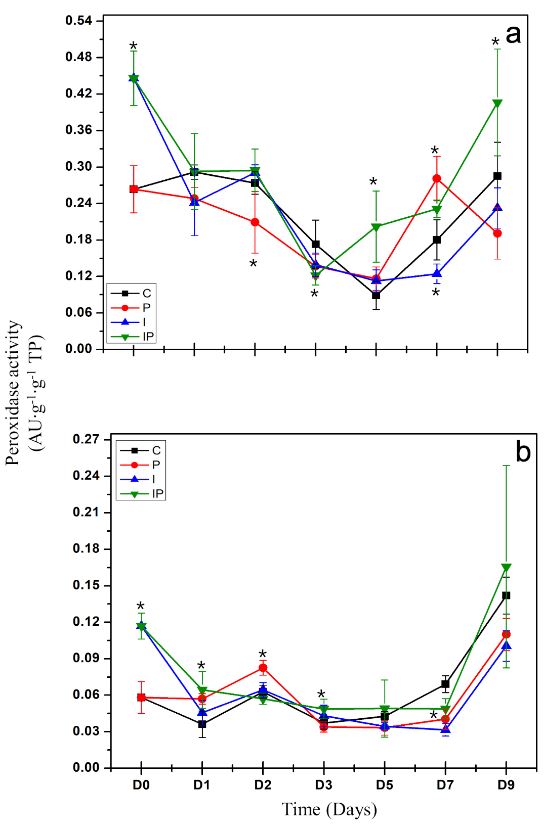
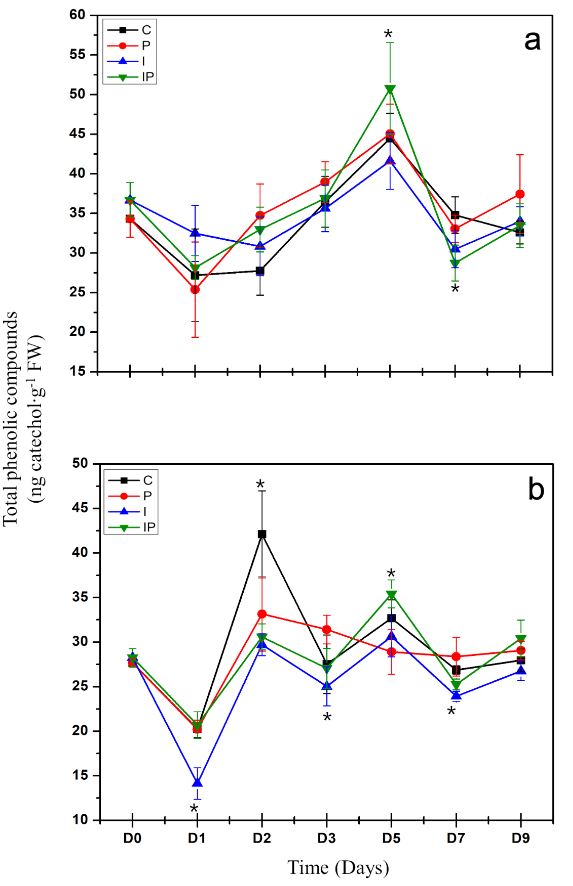
Abstract Background/Objective. Phytophthora capsici is the causal agent of chili wilt. Among the strategies for its control is the use of resistance inducers. Fructans are molecules with interesting biological properties, including the ability to induce resistance mechanisms in some plants. In this work, the protective effect of four concentrations inulin from dahlia tubers on chili infected with P. capsici was evaluated.
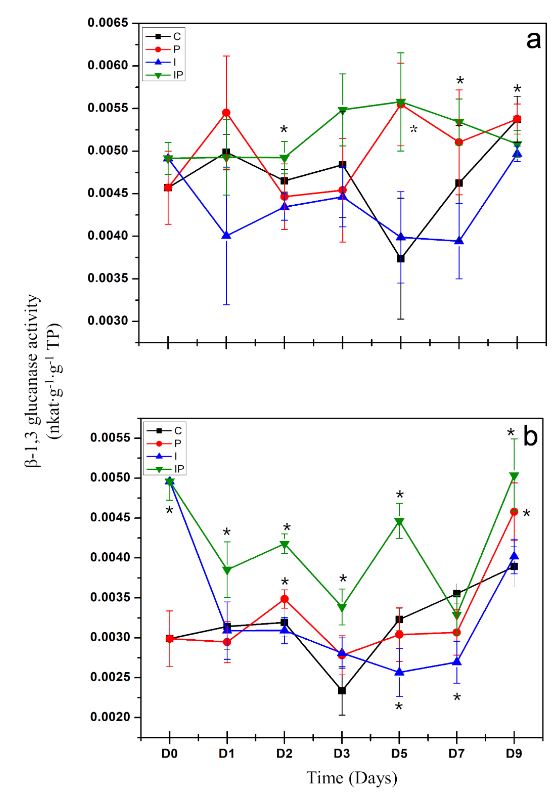
Materials and Methods. The concentration that showed the highest protection was chosen to evaluate the induction of defense response through the enzymatic activity of β-1,3 glucanases, peroxidases and the production of total phenolic compounds.
Results. Inulin showed a protective effect against infection at concentrations of 100 to 300 μM, as symptoms decreased and seedlings showed improved vegetative development. It was observed that inulin at 200 μM concentration was able to induce an effective defense response associated with increased activity of β-1,3 glucanases and peroxidases through a local and systemic response in seedlings. This response was differentiated between seedlings treated with inulin and seedlings infected with P. capsici.
Conclusion. It was concluded that inulin has the ability to protect chili bell pepper from P. capsici by induction of resistance.
Share this link via
Or copy link
byMaría Guadalupe Aguilar Rito, Amaury Martín Arzate Fernández*, Hilda Guadalupe García Núñez, Tomas Héctor Norman Mondragón
Received: 02/October/2023 – Published: 20/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-1
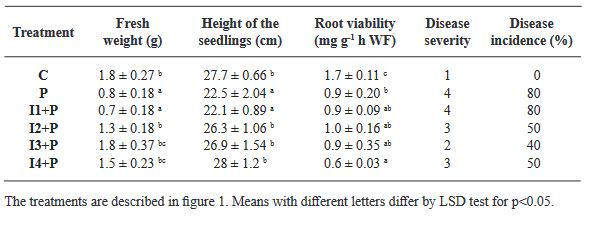
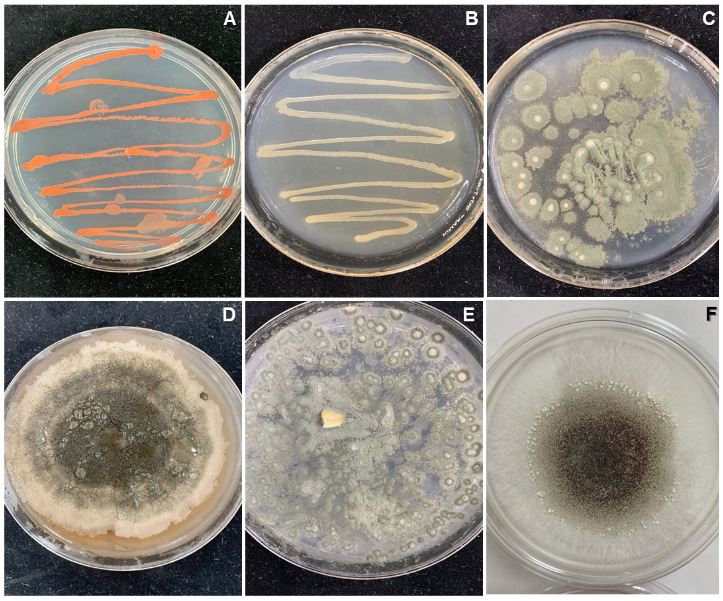
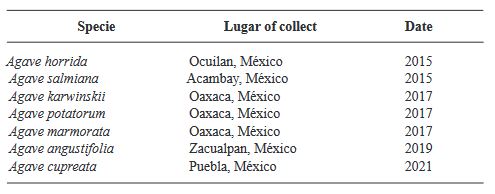
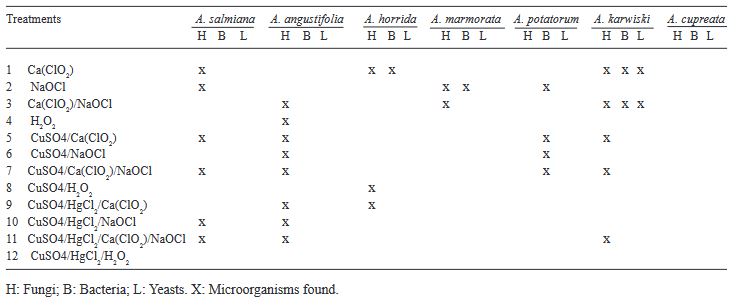
Abstract Background. Disinfection of Agave seeds is a crucial step in in vitro culture to prevent contamination, which can be caused by microorganisms such as bacteria, fungi and viruses that can affect seedling growth and reduce seed germination rate. Therefore, proper seed disinfection is essential to ensure vigorous and healthy plant growth. Objective. Generate an efficient seed disinfection protocol in seven species of Agave; Agave marmorata, A. karwinskii, A. potatorum, A. angustifolia, A. cupreata, A. horrida and A. salmiana to reduce pollution levels.
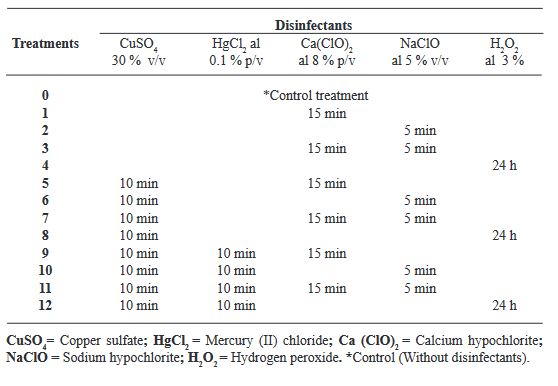
Materials and Methods. A total of 12 disinfection treatments with disinfectants and different combinations were evaluated. The disinfectants used were; 3 % Hydrogen Peroxide for 24 h, Commercial Sodium Hypochlorite 5 % (v/v) for 5 min, Calcium Hypochlorite 8 % (w/v) for 15 min, Copper Sulfate 30 % (v/v) for 10 min, Mercury Chloride II 0.1 % (w/v) for 10 min. Before each treatment was tested, the seeds were pre-washed with liquid soap and subjected to the treatments, Subsequently, they were sown in DM medium and the percentage of germination and contamination for each treatment was evaluated weekly for a period of 30 days. Additionally, the contaminating microorganisms found were identified.
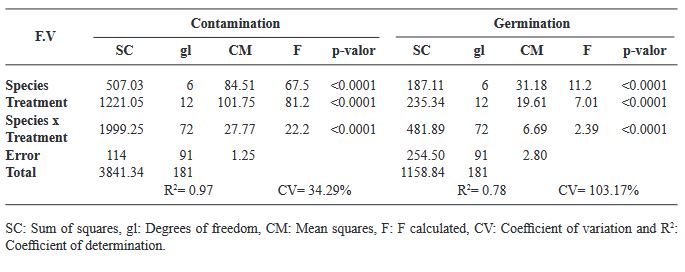
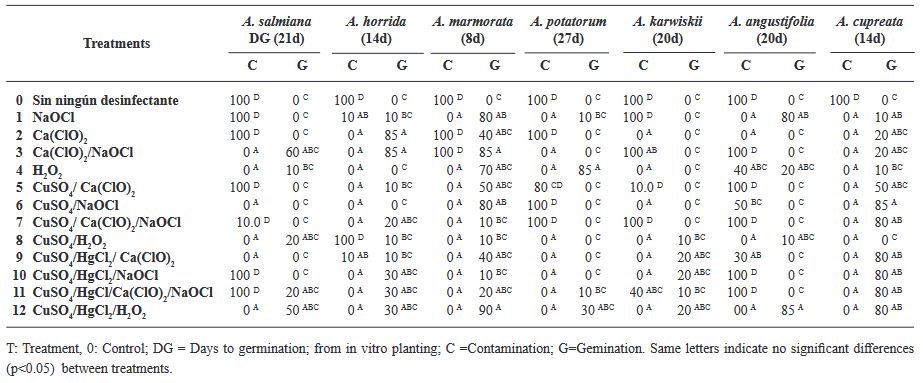
Results. The best treatment for seed disinfection was 30 % copper sulfate (v/v) for 10 min, 0.1 % mercuric chloride II for 10 min and 3 % hydrogen peroxide for 24 h, obtaining 100 % disinfection. Four genera of fungi were identified: Monilinia sp., Aspergillus sp., Penicillium sp., and Alternaria alternata, a bacterium; Bacillus sp., and a yeast, Schizosaccharomyces sp.
Share this link via
Or copy link

Bacillus sp. A8a reduces leaf wilting by Phytophthora and modifies tannin accumulation in avocado
byEdgar Guevara Avendaño, Itzel Anayansi Solís García, Alfonso Méndez Bravo, Fernando Pineda García, Guillermo Angeles Alvarez, Carolina Madero Vega, Sylvia P. Fernández Pavía, Alejandra Mondragón Flores, Frédérique Reverchon*
Received: 17/September/2023 – Published: 08/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2309-2
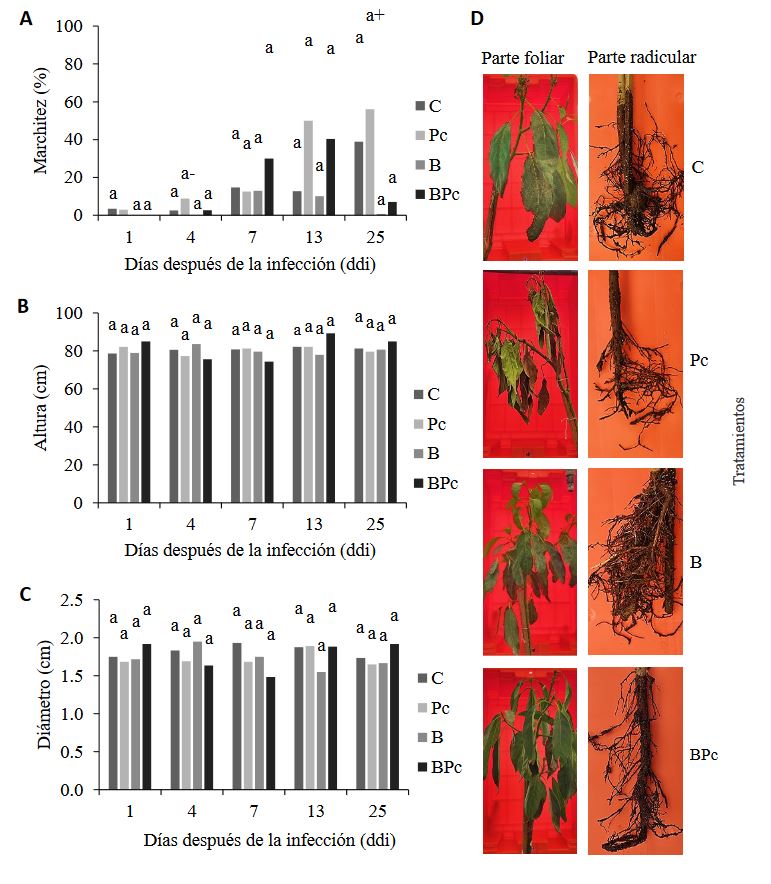
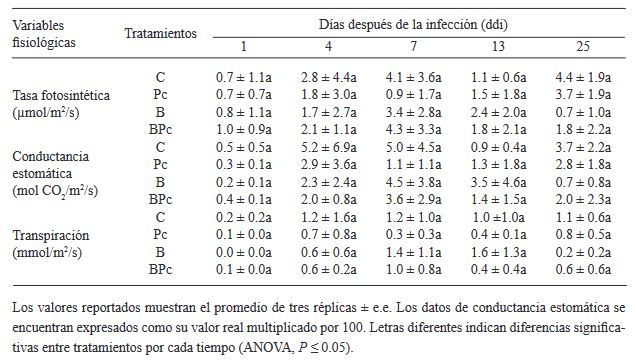
Abstract Background/Objective. The objective was to assess the biocontrol capacity of Bacillus sp. A8a in avocado (Persea americana) plants infected by Phytophthora cinnamomi.
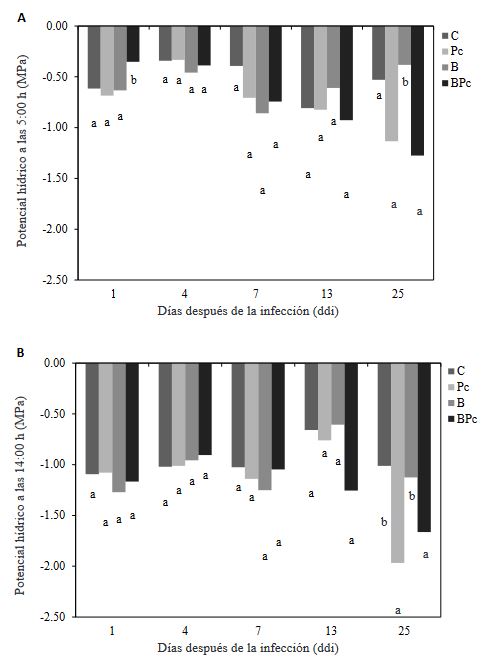
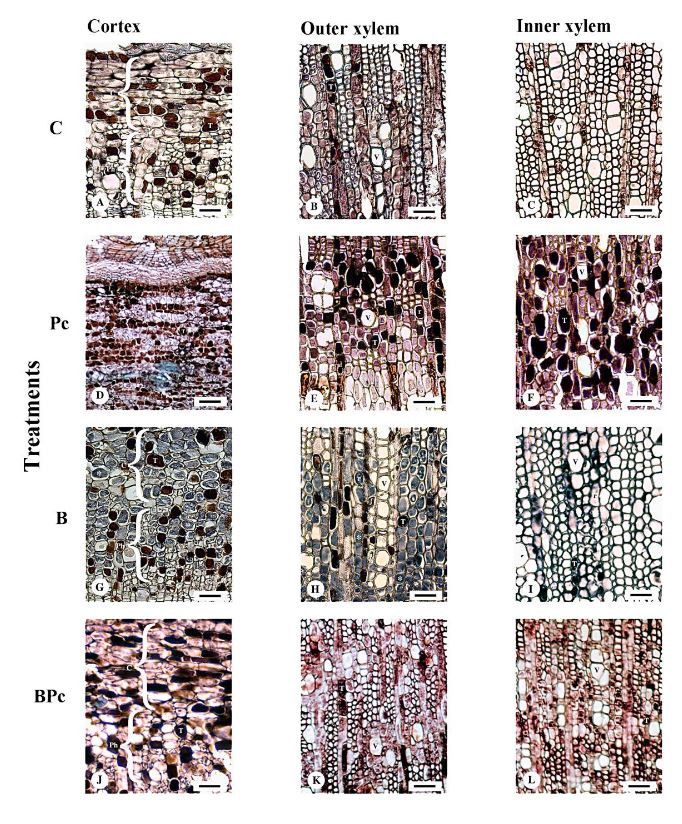
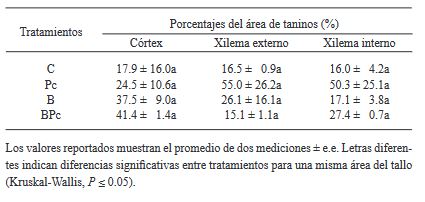
Materials and Methods. A greenhouse experiment was implemented with four treatments: 1) control plants; 2) plants infected with P. cinnamomi; 3) plants inoculated with Bacillus sp. A8a; 4) plants infected with P. cinnamomi and inoculated with Bacillus sp. A8a. We evaluated several morpho physiological variables during the experiment, which lasted 25 days after infection (dai). Moreover, we analyzed tannin density in stems at 25 dai to determine the plant defense response against the disease.
Results. Inoculation with strain A8a reduced wilting symptoms by 49 % at 25 dai, compared with non-inoculated plants. No differences were detected in morpho physiological variables between treatments. However, a greater tannin accumulation was registered in the xylem of infected plants, whilst plants inoculated with strain A8a displayed a larger tannin density in the cortex.
Conclusion. Our results confirm the biocontrol activity of Bacillus sp. A8a in avocado plants and suggest that tannin differential accumulation in the cortex of plants inoculated with the bacteria may contribute to the enhanced tolerance of avocado plants against Phytophthora root rot.
Share this link via
Or copy link
Characterization of endophytic bacteria growth-promoting in potato plants (Solanum tuberosum)
byRosa María Longoria Espinoza*, Cristal Leyva Ruiz, Gloria Margarita Zamudio Aguilasocho, Rubén Félix Gastélum
Received: 02/October/2023 – Published: 23/January/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-4
Abstract Background/Objective. The purpose of this research was to evaluate the in vitro plant growth-promoting activity of endophytic bacteria isolated in tissue from Atlantic variety potato plants from the municipality of Guasave, Sinaloa, Mexico.
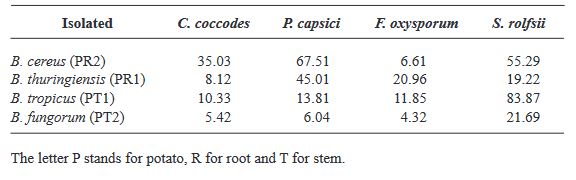
Materials and Methods. The bacterial population was isolated in Lb agar culture medium; two bacterial isolates were obtained from the root and two from the stem, all four Gram positive. The bacterial population of the tissue samples was expressed as (CFU/g-1). The phosphate solubilization capacity, production of chitinases and siderophores were qualitatively evaluated.
Results. Partial sequencing of the 16S rDNA gene was performed, allowing the identification of associated bacterial species within the Firmicutes. 100% of the strains were identified as Bacillus sp. with identities greater than 97%: B. cereus, B. tropicus, B. thuringiensis, B. fungorum. The B. thuringiensis and B. cereus strains showed positive activity in promoting plant growth in vitro through phosphate solubilization, production of chitinases and siderophores. B. cereus and B. tropicus presented inhibitory capacity greater than 50% for Sclerothium rolfsii.
Conclusion. It is relevant to continue research carried out in the laboratory, in order to determine its potential in the field, improving the production of potato crops.
Share this link via
Or copy link