

Share this link via
Or copy link


Article filters
Search Papers
byCélida Aurora Hernández Castillo, Patricia Rivas Valencia*, Leticia Robles Yerena, Mariana Guadalupe Sánchez Alonso, Emiliano Loeza Kuk
Received: 31/May/2024 – Published: 05/November/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-03
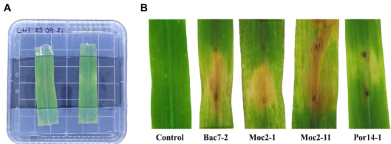
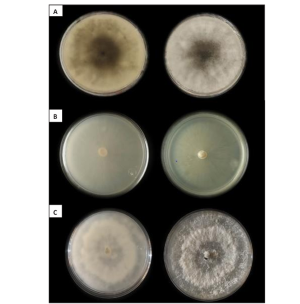
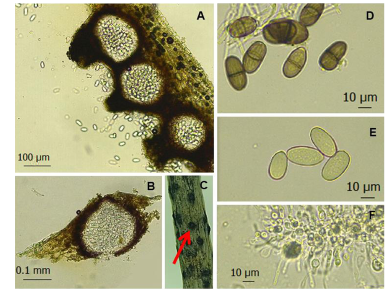
Abstract Background/Objective. In all citrus-producing regions in the world, gummosis is a disease that has caused losses in citrus production. This disease is caused by several pathogens. The objectives were to identify the fungi associated with gummosis in citrus orchards of Plan Chac, Sacalum, Yucatan; and to evaluate chemical and biological alternatives for the control of fungi associated with gummosis.
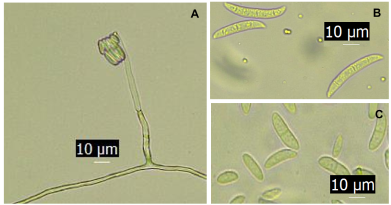
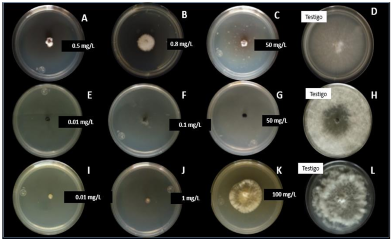
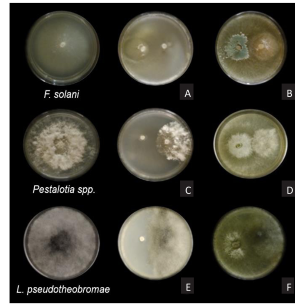
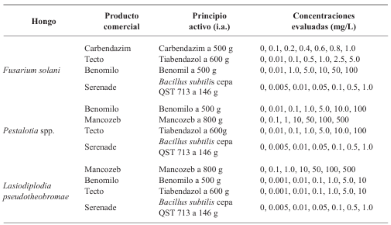
Materials and Methods. From fragments of plant tissue and soil, the associated fungi were isolated. The isolates were identified morphologically in plant tissue as Lasiodiplodia pseudotheobromae and in soil as Fusarium solani and Pestalotia spp. The pathogenicity test determined that L. pseudotheobromae is an agent associated with this disease. The isolates were subjected to in vitro tests with chemical fungicides and antagonist agents.
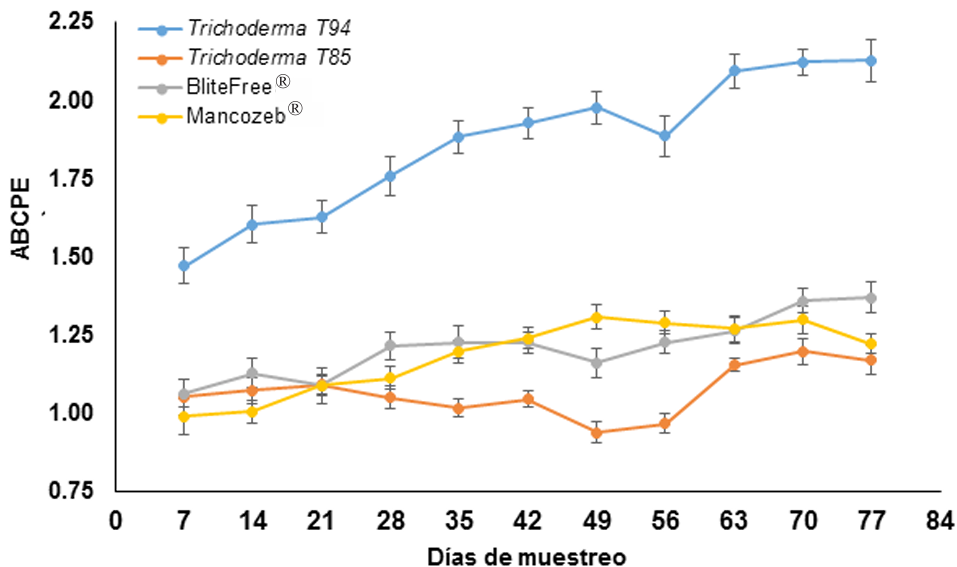
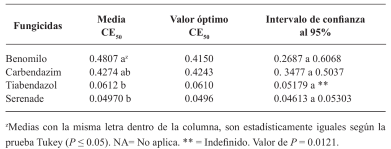
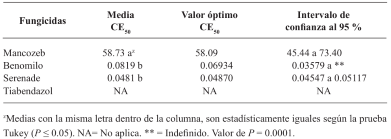
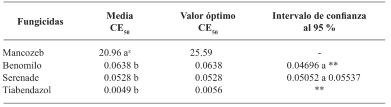
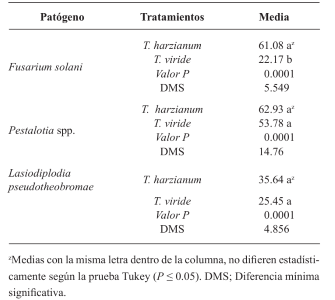
Results. Thiabendazole showed effectiveness for F. solani with an effective concentration to inhibit 50 % of the population (EC50) of 0.0612 mg L-1, with Pestalotia spp. inhibited growth at all concentrations evaluated and for L. pseudotheobromae, it showed an EC50 of 0.0049 mg L-1. In the case of Bacillus subtilis strain QST 713, the growth of F. solani (EC50 0.0496 mg L-1), Pestalotia spp. (EC50 0.0487 mg L-1) and L. pseudotheobromae (EC50 0.0528 mg L-1) decreased. On the other hand, Trichoderma harzianum showed a greater inhibition against F. solani, Pestalotia spp. and L. pseudotheobromae of 61.08, 62.93 and 35.64 %, respectively.
Conclusion. In the management of gummosis in citrus fruits, the use of biological agents such as Trichoderma and B. subtilis can be efficiently included, offering alternatives with less impact on the environment
Share this link via
Or copy link

byLeonardo Miguel Nava Eugenio, Dolores Vargas Álvarez, Eleuterio Campos Hernández, Flaviano Godínez Jaimes, Roxana Reyes Ríos, Mairel Valle de la Paz*, Daniel Perales Rosas
Received: 31/May/2023 – Published: 05/November/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-04
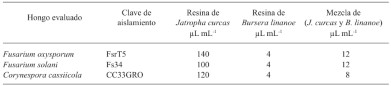
Abstract Antecedents/Objectives. Guerrero is an important producer of roselle (Hibiscus sabdariffa); therefore, the objective was to evaluate the inhibitory effect of resins in vitro using four factors: technique (disc in agar or Kirby Bauer and well in agar), resins (B. linanoe, J. curcas, mixture of both and Thiabendazole), volume (10, 20 and 30 μL) and phytopathogenic fungi (C. cassiicola, F. oxysporum and F. solani) on the diameter of the inhibition halo.
Materials and Methods. Statistical analysis was performed with a completely randomized factorial design with fixed effects to compare the 72 treatments using the Kruskal-Walis test.
Results. All terms were found to be significant, the main effects of technique, resins, volume and fungi on the diameter of the inhibition halo, but also the double, triple and quadruple interactions.
Conclusion. B. linanoe resin showed higher inhibition for C. cassiicola and F. oxysporum, in the two techniques (agar well and agar disc technique or Kirby Bauer technique), this makes it the resin type with the highest biocontrol potential.
Share this link via
Or copy link
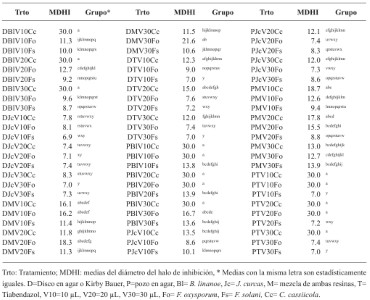
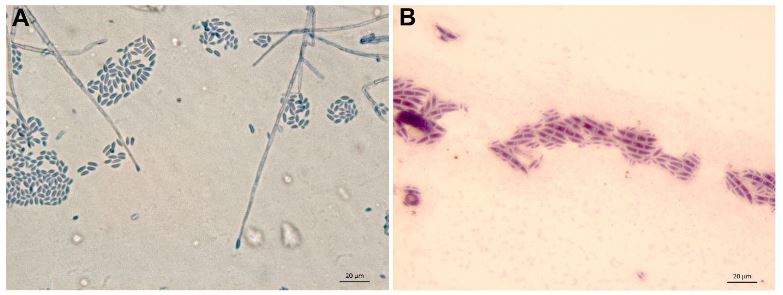
Diversity and taxonomy of Fusarium solani isolated of wilted Agave tequilana var. azul plants
byViviana Montaño Becerrra, Norma Alejandra Mancilla Margalli, Cristina Chávez Sánchez, Martin Eduardo Avila Miranda*
Received: 30/November/2023 – Published: 25/October/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-5
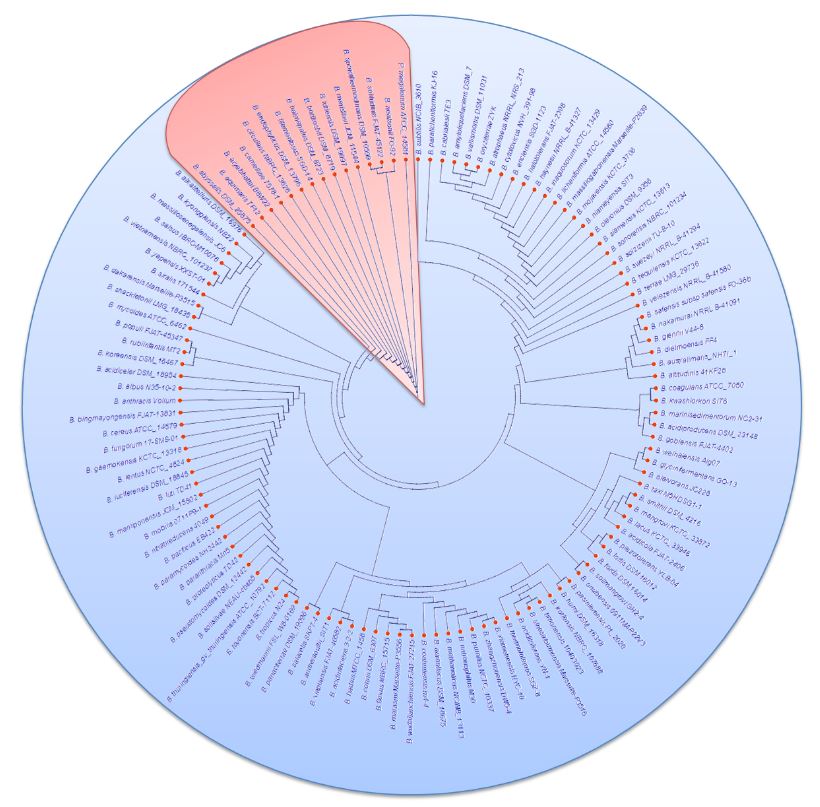
Abstract Background/Objective. The objective of this work was to identify 24 strains of F. solani isolated from agave with wilt, with respect to the new phylogenetic species; determine their molecular similarity to F. solani f. spp.; determine their genetic diversity and their pathogenic capacity in agave, bean and corn.
Materials and Methods. Sequences of the ITS1-5.8S-ITS2 fragment of 24 agave isolates and those of F. solani f. spp., were compared with GenBank and FUSAROID-ID. Amplified 18S rRNA sequences were aligned with sequences reported of F. solani f. spp. phaseoli and batatas, defining the presence of introns. Genetic diversity was determined with the DNA RepPCR marker. Representative strains were tested against agave, bean and maize seedlings, evaluating their pathogenicity as root rot severity.
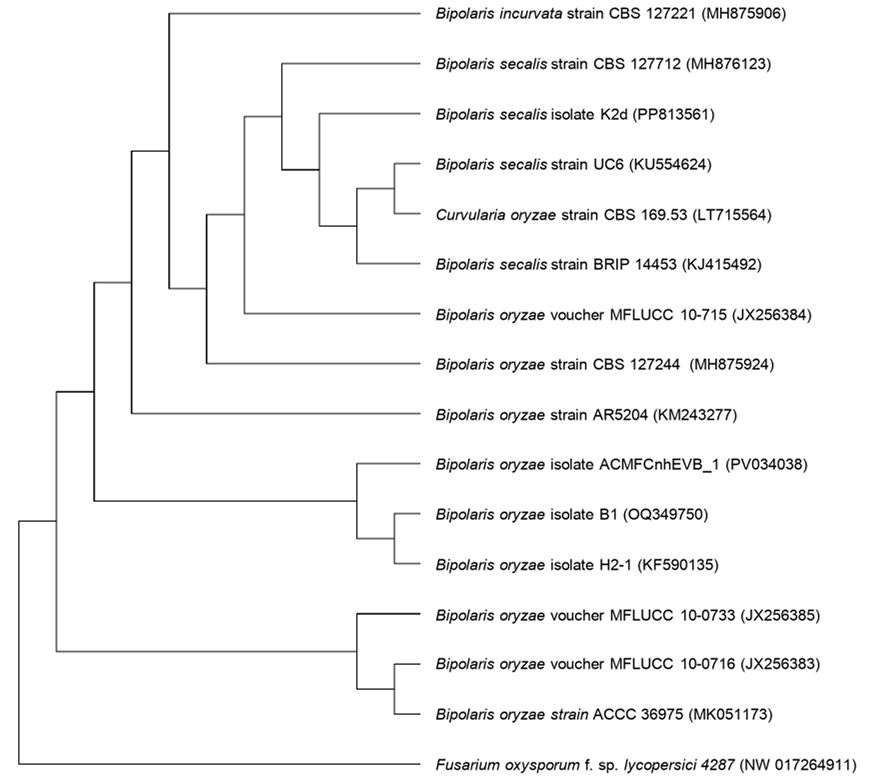
Results. Isolates morphologically identified as F. solani, GenBank placed them as F. solani or included in the FSSC, three strains were identified as Xenoacremonium sp. FUSAROID-ID defined that the sequences of F. solani were highly similar to those of Neocosmospora martii, N. pseudoradicicola, N. solani and N. falciformis. The ITS1-5.8S-ITS2 sequences and absence of introns in its SSU indicated that none is F. solani f. sp. phaseoli. Isolates obtained from agave were pathogenic to A. tequilana and a criollo corn cv, but not to Fusarium-resistant corn. No agave isolates were pathogenic to beans.
Conclusions. Four phylogenetic species of FSSC cause root rot in agave; F. solani isolates from agave did not affect Fusarium-resistant corn. It is safe to intercrop beans in agave.
Share this link via
Or copy link

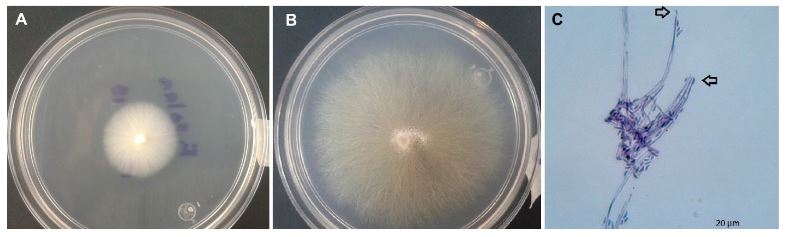
byDiana Angelina Urias Lugo, Octavio Ernesto Martínez Ereva, Cecilia de Los Ángeles Romero Urías, Carlos Ramiro Ibarra Sarmiento, Sylvia Adriana Estrada Díaz, Rubén Félix Gastélum, Karla Yeriana Leyva Madrigal, Guadalupe Arlene Mora Romero*
Received: 24/May/2024 – Published: 24/October/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-01
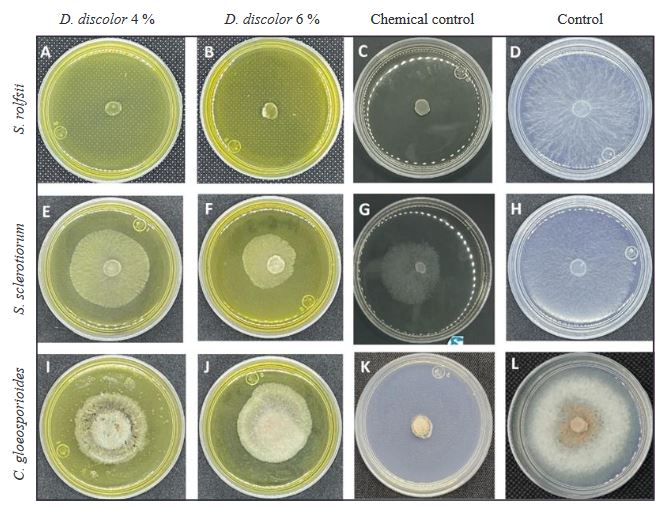
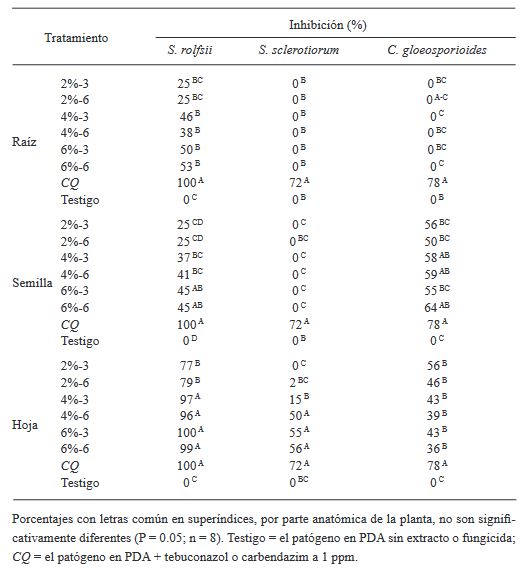
Abstract Background/Objective. The present work reports the in vitro effect of aqueous extracts (2, 4 and 6% w/v) of root, seed and leaf of Datura discolor obtained in two times (3 and 6 minutes) at High Pressure Processing, against Sclerotium rolfsii, Sclerotinia sclerotiorum and Colletotrichum gloeosporioides.
Materials and Methods. Extracts of roots, seeds and leaves of D. discolor were prepared in a 1:10 w/v ratio with distilled water. Two continuous treatments of high pressure (600 MPa) with pressure maintained for 3 min and another with pressure (600 MPa) maintained for 6 min were considered. The extracts were evaluated against S. rolfsii, S. sclerotiorum and C. gloeosporioides. The experiments were performed in Petri dishes with PDA medium. The efficiency of the extracts was evaluated by obtaining the percentage of inhibition.
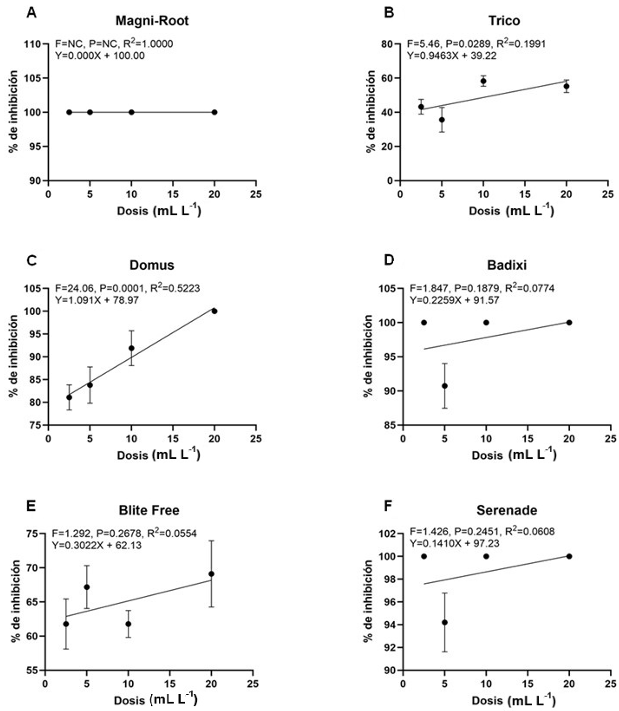
Results. The results show variable percentages of inhibition of the extracts in the different anatomical parts of the plant and concentrations; The leaf extracts at 6%, regardless of the extraction time, show effectiveness against the three pathogens, with inhibition of 99 and 100%, 55 and 56%, and 43 and 36% for S. rolfsii, S. sclerotiorum and C. gloeosporioides at 3 and 6 minutes respectively.
Conclusion. The effectiveness of leaf extract at 6%, six months after its preparation, is similar to the observed with fresh extracts. These results pave the way for future research focused on the sustainable management of phytopathogens. Studies on the biological effectiveness of the extracts in the greenhouse and field are suggested.
Share this link via
Or copy link
Tobamovirus fructirugosum an emerging disease: review and current situation in Mexico
byUbilfrido Vásquez Gutiérrez, Juan Carlos Delgado Ortiz*, Gustavo Alberto Frías Treviño, Luis Alberto Aguirre Uribe, Alberto Flores Olivas
Received: 28/January/2024 – Published: 15/October/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2401-7
Abstract Background/Objective. Tobamovirus fructirugosum species (ToBRFV) is considered a worldwide quarantine pest that limits the production of Solanum lycopersicum and Capsicum annum, currently present in three countries of the American continent. The objective of this work was to deepen in the genetic variability of ToBRFV with respect to the different isolates, the physico-molecular and symptomatic characterization, the traditional and more current methods implemented for diagnosis, the range of virus reservoir hosts, and the epidemiology. Results. ToBRFV was generated from a mutation resulting from genetic recombination with TMV, considered the main progenitor and ToMMV secondary progenitor. Phylogenetic analyses report the existence of five clades with respect to the genetic diversity of ToBRFV. The first primers for detection were designed in 2015 that encode replication, movement and capsid proteins. Serological methods can be used for preventive diagnosis, while molecular and NGS can confirm virus infection even at low concentrations in the plant. Sixteen weed families and host crops are reported from 47 countries. To achieve an effective strategy, it is necessary to reduce inoculum sources, develop compounds that inhibit mechanical transmission and develop tolerant genotypes. Conclusion. ToBRFV is distributed nationally and represents a phytosanitary risk for Mexico; the exhaustive analysis of the study of diagnostic techniques, host range, dissemination, epidemiology and control strategies, contributes to the knowledge of ToBRFV.
Share this link via
Or copy link



byMaria Magdalena Rivera Salas, José Basilio Heredia, Juan Manuel Tovar Pedraza, Cesar San Martín Hernández, José Benigno Valdez Torres, Isabel Cruz Lachica, Raymundo Saúl García Estrada*
Received: 22/May/2024 – Published: 23/August/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2405-11
Abstract Background/Objective. Species of Bacillus are currently gaining interest because its ability to produce secondary metabolites with antifungal properties against various plant pathogenic fungi. The objective of this study was to optimize fermentation temperature and time for antifungal extract production by Bacillus amyloliquefaciens B17 and to verify its activity against plant pathogenic fungi Gilbertella persicaria, Choanephora cucurbitarum, Colletotrichum asianum, and Botrytis cinerea.
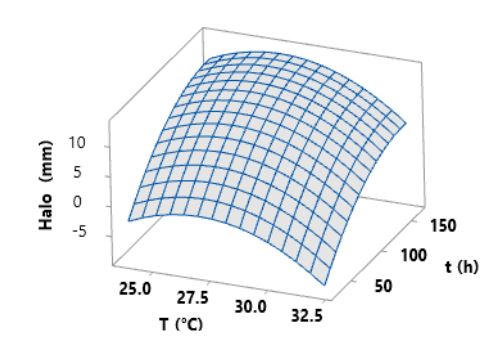
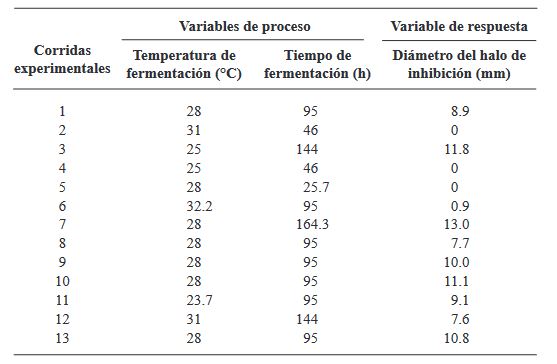
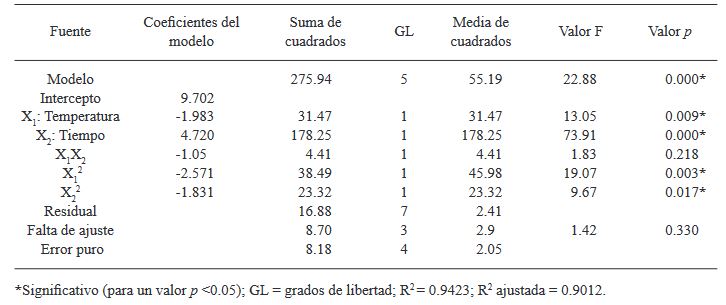
Materials and Methods. A central composite design (CCD) with two factors and five levels (fermentation temperature: 23.7, 25, 28, 31, and 32.2 °C and fermentation time: 25, 46, 95, 144, and 164.3 h) was used. Thirteen combinations of temperature and fermentation time were randomly performed. The thirteen crude extracts of B. amyloliquefaciens B17 were obtained from the cell-free fermentation broth by acid precipitation followed by alkaline solubilization. The response variable was the diameter of the inhibition halos generated by placing drops of the different crude extracts onto the medium already inoculated with a suspension of Gilbertella persicaria spores.
Results. The optimal conditions for the production of the extract with the greatest antifungal activity in B. amyloliquefaciens B17 were 26.8 °C and 158.6 h.
Conclusion. The optimized crude extract from B. amyloliquefaciens B17 exhibited a strong ability to inhibit mycelial growth and spore germination of Gilbertella persicaria, Choanephora cucurbitarum, Colletotrichum asianum, and Botrytis cinerea.
Share this link via
Or copy link
byErick Ortega Piña, Daniel Leobardo Ochoa Martínez*, Reyna Isabel Rojas Martínez, Alfredo Díaz Lara
Received: 15/February/2024 – Published: 12/August/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2402-8
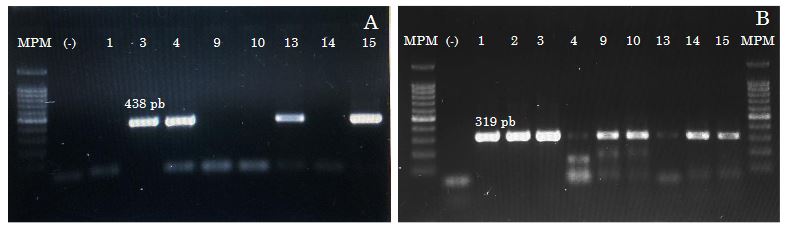
Abstract Background/Objective. In broad bean (Vicia faba) crops established in Montecillo, Texcoco, State of Mexico, 100% of plants were observed with viral symptoms consisting of mosaic, mottling, leaf curling, wilting and a notable decrease in plant development. These symptoms led to a significant reduction in seed yield and quality. In this context, the present study was carried out with the objective of identifying the viral species associated with these symptoms.
Materials and Methods. The experimental strategy consisted of extraction of total RNA from leaves showing the aforementioned symptoms, followed by nextgeneration sequencing.
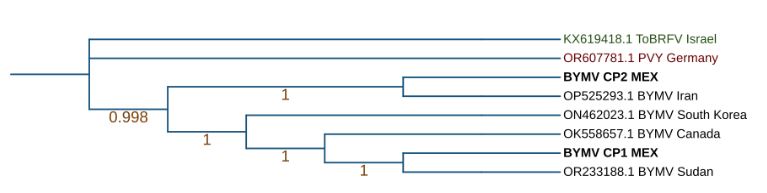
Results. In the results, the complete genomes of Orthotospovirus impatiensnecromaculae (before impatiens necrotic spot virus) and two isolates of the Potyvirus phaseoluteum (before bean yellow mosaic virus) were obtained. The phylogenetic analysis revealed that the isolate of O. impatiensnecromaculae (INSV CPMEX) shows significant divergences from those previously reported in other plant species. On the other hand, the two P. phaseoluteum isolates (BYMV CP1MEX and BYMV CP2MEX) proved to be different from each other, being related to isolates reported in Sudan and Iran.
Conclusion. This result suggests considerable genetic diversity among the viruses associated with viral symptoms in faba bean crops in the region, which underlines the importance of accurate identification for the management and control of these viral infections.
Share this link via
Or copy link

byYeison David López Galé*, Mauricio Fernando Martínez, Lizeth Paola Palacios Joya, Nubia Murcia Riaño, Mario Augusto García Dávila
Received: 22/November/2023 – Published: 08/August/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2311-1
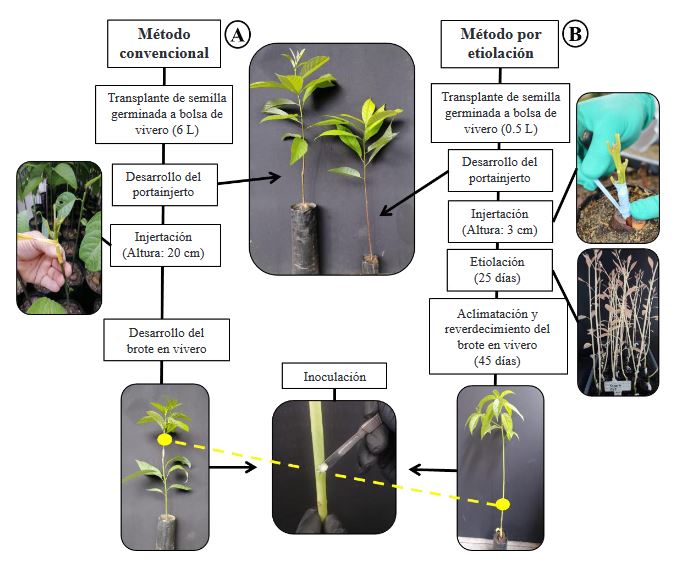
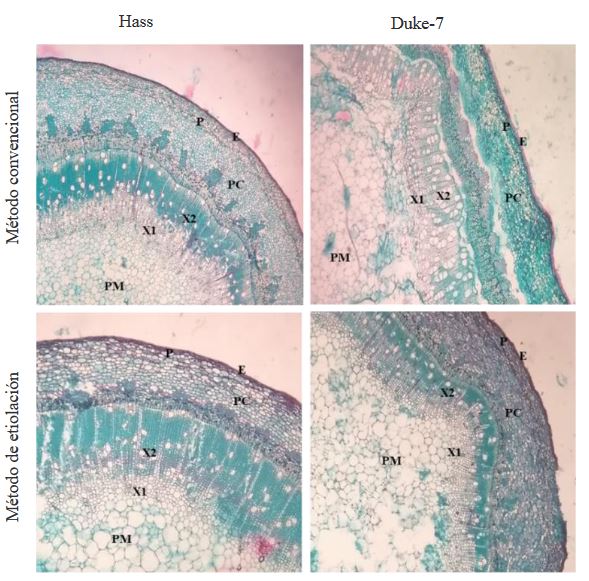
Abstract Background/Objective. The level of resistance to Phytophthora cinnamomi in avocado germplasm can be evaluated indirectly through inoculation of the pathogen by wounding the stem. The objective of this work was to compare the conventional graft development method and the etiolated graft method to determine levels of indirect resistance to P. cinnamomi through the stem wound inoculation technique.
Materials and Methods. In the study, three isolates of P. cinnamomi and two avocado genotypes with different levels of resistance to the pathogen were used, Duke-7 (medium resistant) and Hass (susceptible). Clonal multiplication of the genotypes was carried out with buds grafted on rootstocks propagated by Antillean avocado seeds. Inoculation was performed on the shoot at a height of 8 cm and the growth of the lesions was measured for 24 days. With the data, the Area Under the Disease Progress Curve (AUDPC) and the Coefficients Variation (CV) are calculated. The information was analyzed with a completely randomized experimental design with a 2*2*3 factorial arrangement (Method*Genotype*Isolation).
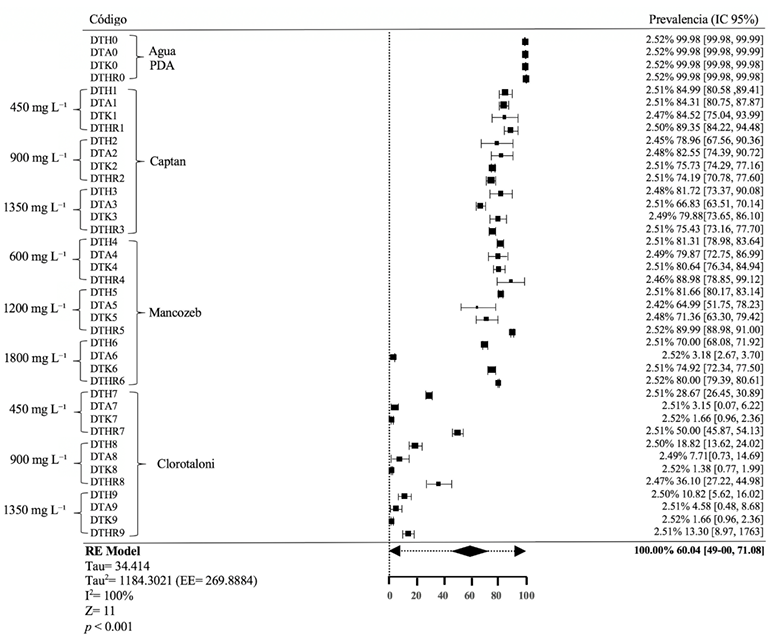
Results. The analysis of variance for the AUDPC showed no differences between methods (p=0.1881); However, there were differences between genotypes, isolates and between the genotype*method and genotype*isolation interactions (p≤0.05). With the conventional method, the development of the outbreaks was late (141-159 days) and the size of the lesions was highly variable (CV=38.9-64.4%), being able to discriminate partial degrees of aggressiveness between isolates, but not levels of resistance between genotypes. The etiolated and greened shoots in the nursery, on the contrary, presented rapid growth (101-107 days) and greater uniformity in the lesions generated by the pathogen (CV=11.1-24.2%).
Conclusion. The development of etiolated shoots in avocado grafts is proposed as a rapid alternative method that can guarantee greater uniformity in the development of lesions within the experimental units of a treatment, thus achieving greater reliability when evaluating and selecting preliminarily. avocado genotypes with indirect resistance attributes to P. cinnamomi.
Share this link via
Or copy link
Mixed viral infections in vegetable crops: biochemical and molecular aspects
byMario Sánchez Sánchez, Irasema Vargas Arispuro, Juan Manuel Tovar Pedraza, Cristóbal González Pérez Pedraza, Emmanuel Aispuro Hernández, Eber Addí Quintana Obregón, Miguel Ángel Martínez Téllez*
Received: 05/April/2024 – Published: 06/August/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-3
Abstract Mixed viral infections refer to the coinfection of two or more viruses in the plant, which regularly lead to exacerbated symptoms on leaves and fruits. The dynamics of coinfections may follow either a synergistic, antagonistic, or neutral interaction that impacts the severity of the symptoms and infection. Mixed viral infections occur due to the convergence of fundamental characteristics reviewed in this manuscript. The virus‒host plant interrelationship influences the establishment and spread patterns of mixed viral infections. Attention should be drawn to potential changes in the dynamics of transmission and prevalence of plant viral diseases due to the effect of anthropogenic and natural alterations to complex agroecological systems or their components, including hosts, reservoirs, vectors, ecological niches, and the emergence of new virus strains
Share this link via
Or copy link
![Figure 1. Symptom transition in the lower, middle, and upper leaves of tomato plants agroinoculated with the infectious cloned DNA-A and DNA-B components of isolates of TYLCV from Costa Rica (ToYMoV-[CR:Gre:GR1:90]) and Tomato leaf curl Sinaloa virus (ToLCSiV-[CR:Lib:L1:02]) from Costa Rica and the infectious clone of the genomic DNA of TYLCV from the Dominican Republic (TYLCV-[DO]) individually or in all combinations. (A) TYLCV; (B) non-inoculated tomato plant; (C) ToYMoV and ToLCSiV; (D) ToYMoV and TYLCV; (E) ToLCSiV and TYLCV; (F) ToYMoV, ToLCSiV and TYLCV. Plants were photographed 21 d after agroinoculation (with permission of Maliano <em>et al.,</em> 2022)](img/RMF/Volumenes/NumNormales/V4232024/RMF2404-3/Figure1.jpg)
byRubén Félix Gastélum*, Gabriel Herrera Rodríguez, Norma Ávila Alistac, Elizabeth León
Received: 22/April/2024 – Published: 06/August/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-6
Abstract White mold (Sclerotinia sclerotiorum) is the main disease of bean and potato in Sinaloa. In the present review, the symptoms and signs of the disease as well as cultural and morphological characteristics of the teleomorph of the pathogen, its ecology and the epidemiology of the disease are addressed. The implementation of a prediction system which includes the carpogenic germination of the sclerotia and the phenology of both bean and potato for the management of the disease is described. This system considers soil temperature ranging from 13 to 19 °C a at depth of 2.5 cm in the soil and the flowering stage in both bean and potato to do the first spray application of synthetic fungicide to prevent the disease. in vitro studies indicated that Trichoderma harzianum, T. viride and T. atroviride reduced mycelial growth rate of S. sclerotiorum. The same antagonistic species exerted control of white mold under field conditions, where an increment of 40% of yield was observed in the treated plots, with respect to those treated with fungicide fluazinam. Future lines of research focusing on the ecology of the pathogen and management of the disease including the antagonistic fungi in the prediction system are suggested.
Share this link via
Or copy link
