


Share this link via
Or copy link


Article filters
Search Papers
byAna María López López, Juan Manuel Tovar Pedraza, Josefina León Félix, Raúl Allende Molar, Nelson Bernardi Lima, Isidro Márquez Zequera, Raymundo Saúl García Estrada*
Received: 27/September/2023 – Published: 13/February/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2309-5
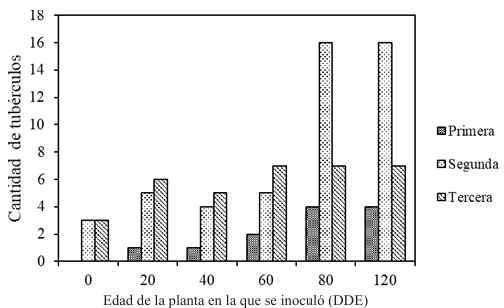
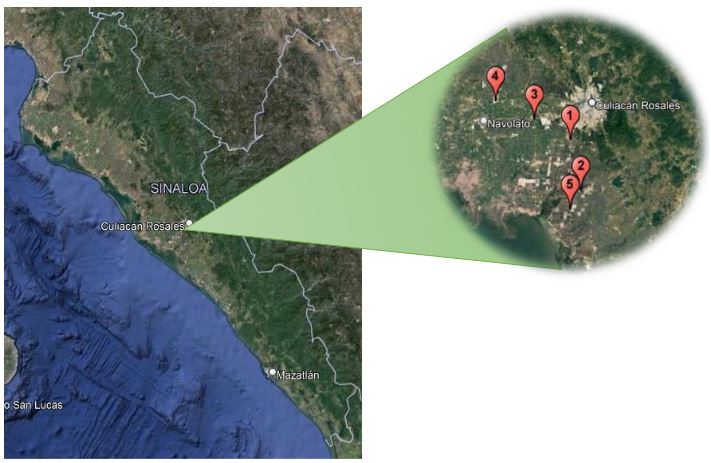
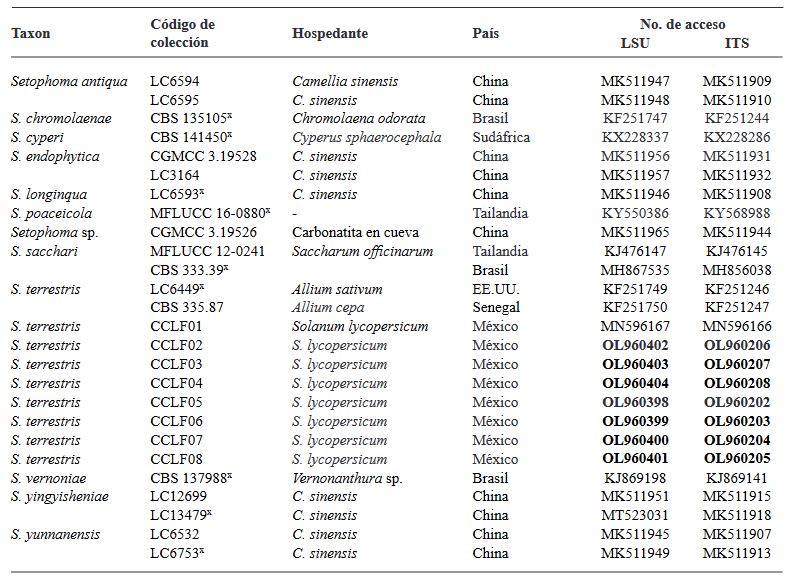
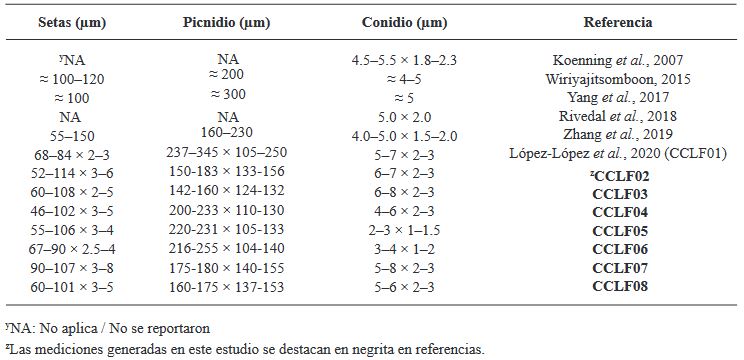
Abstract Background/Objective. Tomato (Solanum lycopersicum) is one of Mexico’s main crops. In the years 2017 and 2018, symptoms of corky and pink roots were observed with an incidence of 10 to 20% in Culiacan, Sinaloa, Mexico. In the foliage, plants presented a generalized chlorosis, with stunted growth and senescence in the leaves. In the roots, brown and pink lesions were formed, as well as a corky texture. The objective of this study was to morphologically and molecularly characterize fungal isolates associated to corky and pink root in tomato orchards in Culiacan, Sinaloa, as well as to evaluate their pathogenicity.
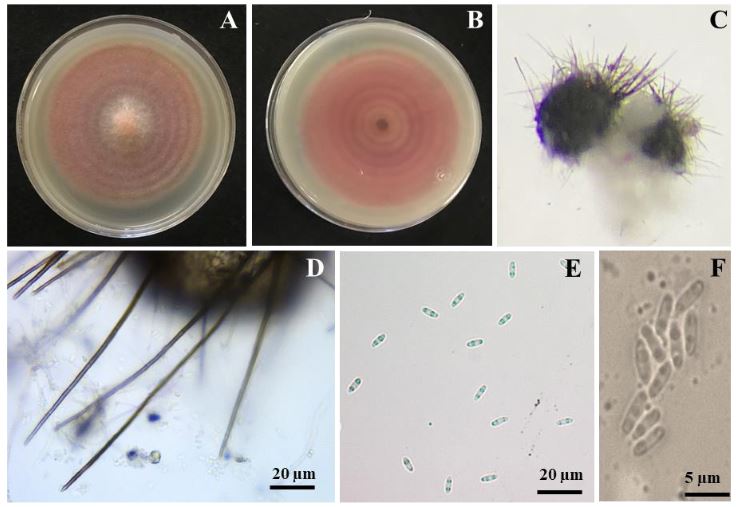
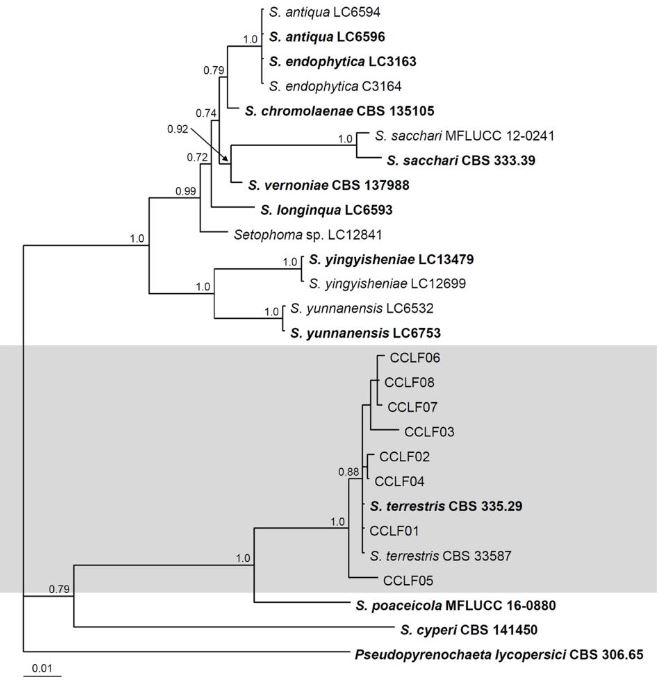
Materials and Methods. Monoconidial isolates were obtained and they were identified as Setophoma terrestris, based on their morphological characteristics. To confirm the identity, the area of the internal transcribed spacers (ITS) of the rDNA was amplified and sequenced, along with a fragment of the gene 28S of the rRNA (LSU).
Results. Using the sequences obtained, a phylogenetic tree was created using the Bayesian Inference and it was found that the sequences were grouped with the ex–type sequences of Setophoma terrestris. The pathogenicity of the isolates was verified by inoculating mycelial discs into the root of 10 one-month-old tomato seedlings. The roots of the seedlings inoculated with PDA discs without mycelium served as a control. Thirty days after inoculation, corky and pink root symptoms appeared, whereas the roots of control plants remained healthy
Conclusion. According to the morphological characterization, the molecular identification and the pathogenicity tests, Setophoma terrestris was confirmed to be the causal agent of corky and pink root in agricultural tomato orchards in Culiacan, Sinaloa.
Share this link via
Or copy link


Genus Orthotospovirus in Costa Rica: A Central American case
byMauricio Montero Astúa*, Natasha Dejuk Protti, David Bermúdez Gómez, Elena Vásquez Céspedes, Laura Garita Salazar, Federico J. Albertazzi, Scott Adkins, Lisela Moreira Carmona
Received: 23/August/2023 – Published: 28/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-6
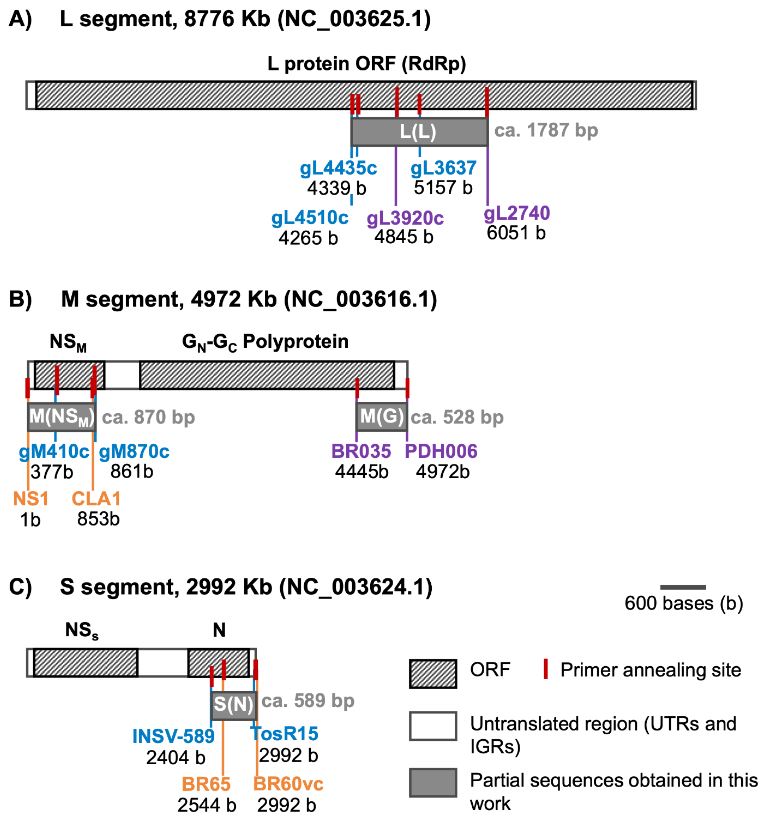
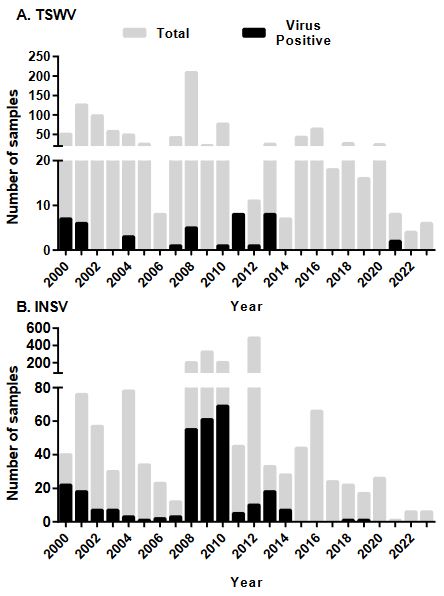
Abstract Background/Objective. The Orthotospovirus genus encompasses a range of economically significant and emerging plant viruses that affect a variety of crops globally. While the prevalence and characteristics of these phytopathogenic viruses are extensively documented in North and South America, their presence in Central America remains comparatively underexplored. This study focuses on Costa Rica, strategically positioned at the nexus of North and South America, to enhance our understanding of Orthotospovirus in this region.
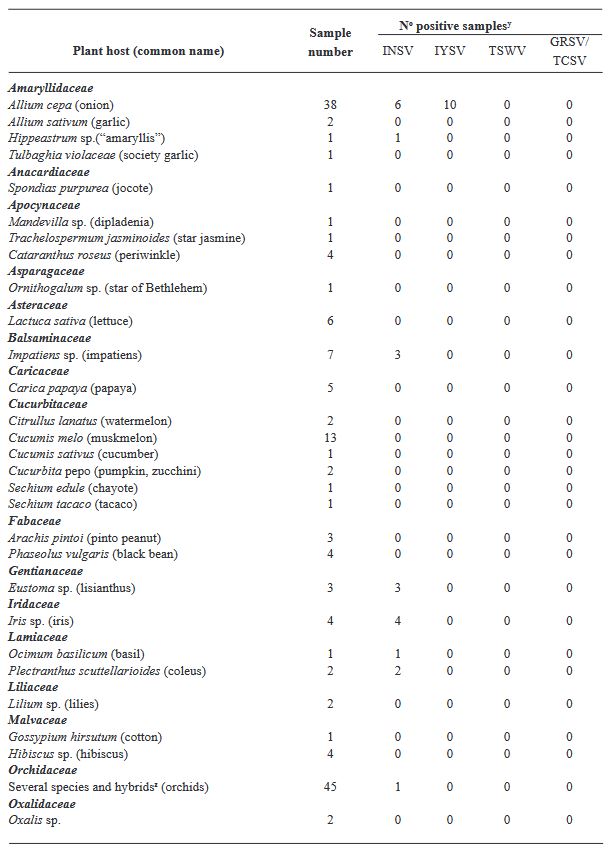
Materials and Methods. We analyzed 295 plant samples using enzyme-linked immunosorbent assay (ELISA) to test for the presence of INSV, IYSV, TSWV, and the GRSV/TCSV serogroup. Additionally, a subset (20 samples) underwent further scrutiny through reverse transcription-polymerase chain reaction (RT-PCR) employing both universal and species-specific primers.
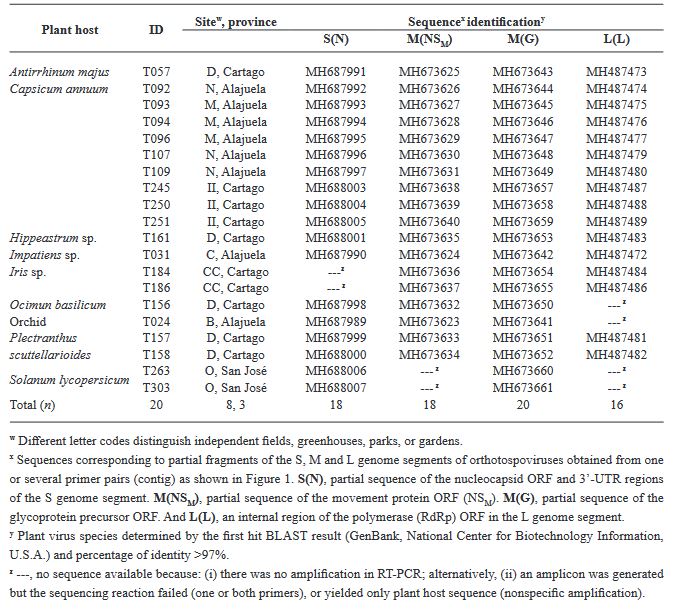
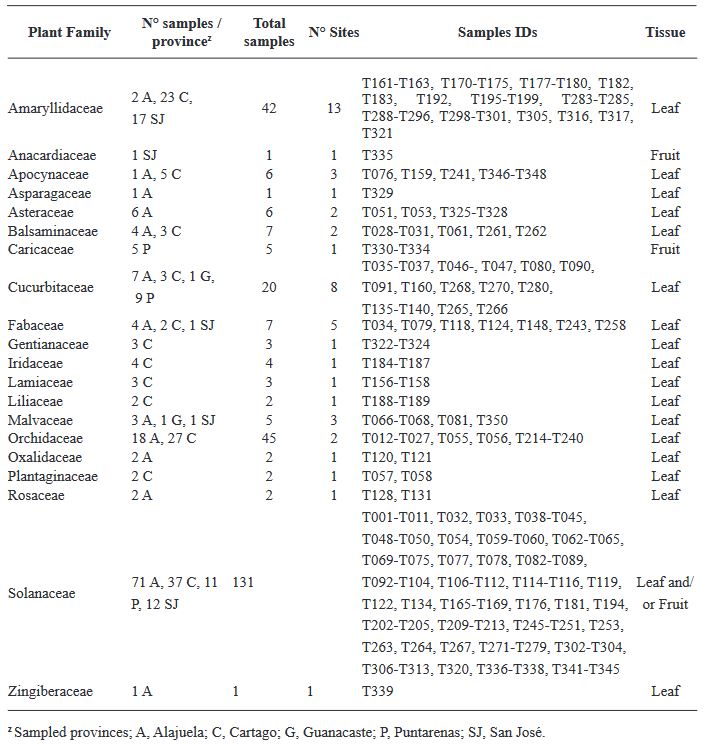
Results. Our ELISA results indicated the absence of TSWV and the GRSV/TCSV serogroup. However, the presence of INSV in Costa Rica was substantiated through ELISA, RT-PCR, and partial sequencing, revealing its prevalence in both open-field and greenhouse environments. Despite previous diagnostic reports suggesting the presence of TSWV in Costa Rica, our study did not detect this virus. RT-PCR analysis with degenerate primers also found no evidence of other Orthotospovirus species in our samples. The identification of a dominant INSV haplotype, along with three additional variants, suggests the likelihood of at least two independent virus introductions into the region.
Conclusion. These findings underscore the necessity for more comprehensive surveys and research on Orthotospoviruses in Central America to better understand their epidemiology and impact on agriculture.
Share this link via
Or copy link

![Figure 4. Haplotype Network Graph of Impatiens Necrotic Spot Virus. This graph is a haplotype network inferred using the TCS method, based on a 261 bp sequence alignment of the nucleocapsid protein ORF (S[N] segment) of Impatiens necrotic spot virus. The network includes data from 81 isolates, visually representing the genetic relationships and diversity among the different haplotypes of the virus.](img/RMF/Volumenes/NumEspeciales/VE4142023/RMF2023-6/Figure4.jpg)
![Figure 5. Comprehensive phylogenetic analysis of impatiens necrotic spot virus isolates. This figure illustrates a phylogenetic tree constructed from an alignment of concatenated sequences (2945 positions) encompassing regions of the nucleocapsid [S(N)], glycoprotein precursor [M(G)], movement protein [M(NSM)], and viral polymerase [L(L)] ORFs. The tree includes isolates from various countries, identified by ISO 3166-1 three- letter codes: CHN (China), ITA (Italy), KOR (South Korea), and USA (United States of America). For Costa Rican isolates, additional distinct letter codes in parenthesis signify separate geographic locations. The analysis, executed in MEGA X, utilized the Maximum Likelihood method with a Tamura-3-parameter model and a gamma-distributed rate of variation in nucleotides (+G), involving 2000 permutations. The scale bar indicates the number of nucleotide substitutions per site.](img/RMF/Volumenes/NumEspeciales/VE4142023/RMF2023-6/Figure5.jpg)
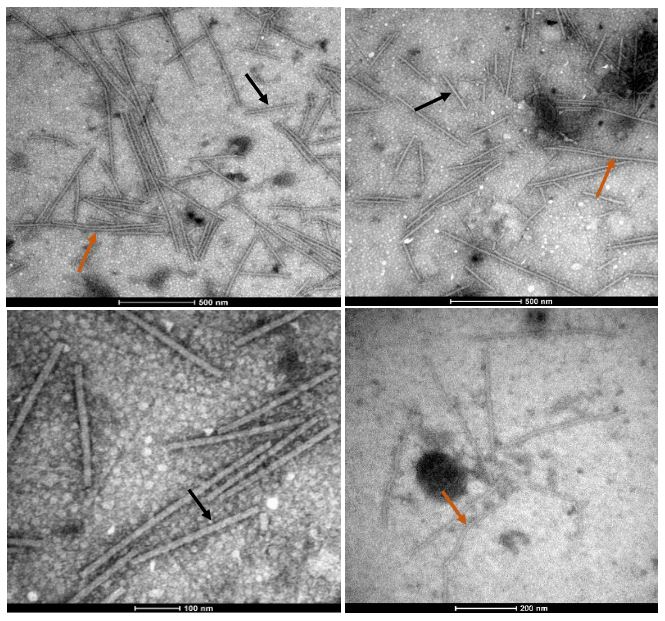
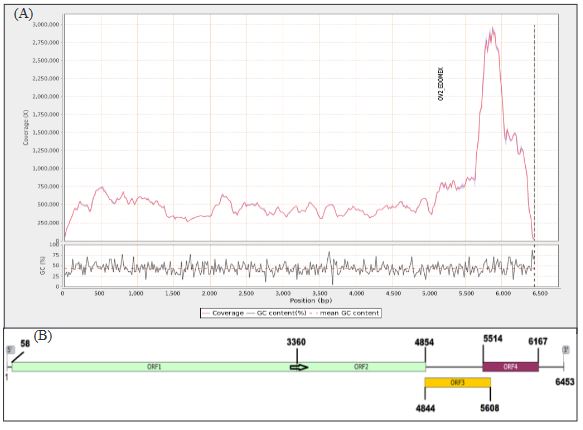
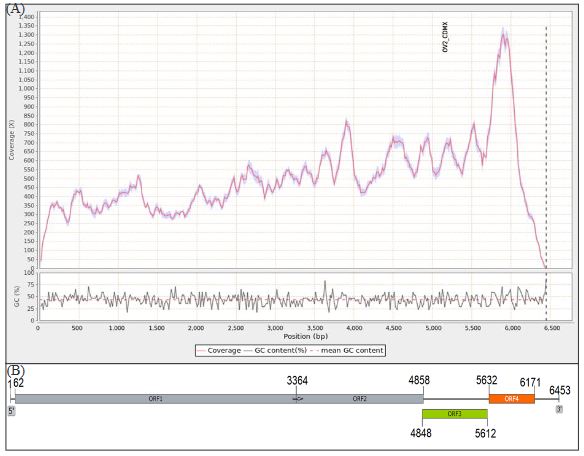
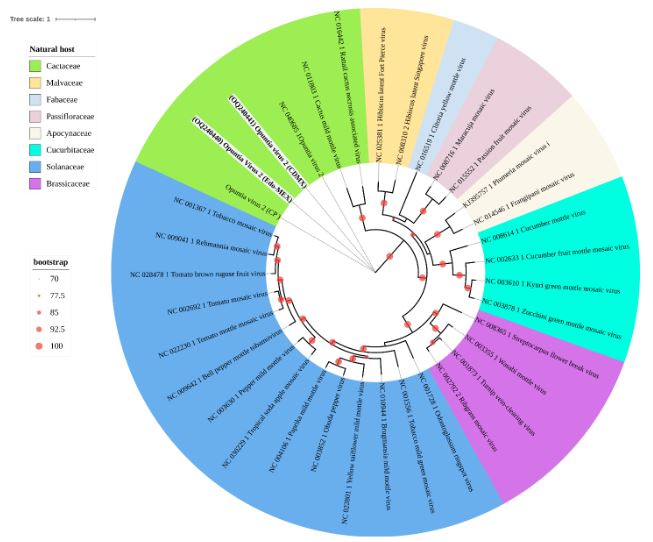
Virome of the vegetable prickly pear cactus in the central zone of Mexico
byCandelario Ortega Acosta, Daniel L. Ochoa Martínez*, Reyna I. Rojas Martínez, Cristian Nava Díaz, Rodrigo A. Valverde
Received: 31/July/2023 – Published: 27/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-2
Abstract Background/Objective. In this study, the ability of high-throughput sequencing (HTS) to detect viruses in vegetable prickly pear cactus was exploited.
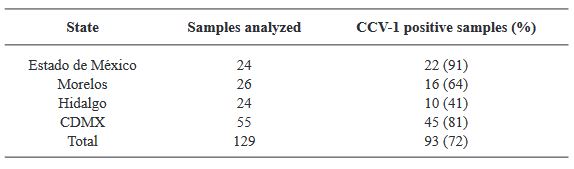
Materials and Methods. Samples from State of Mexico (EDMX), Hidalgo, and Morelos, as well as Mexico City (CDMX), were analyzed.
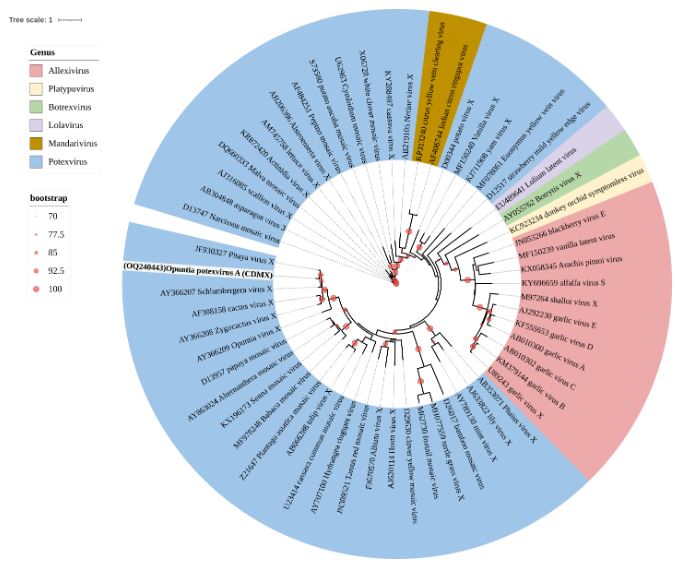
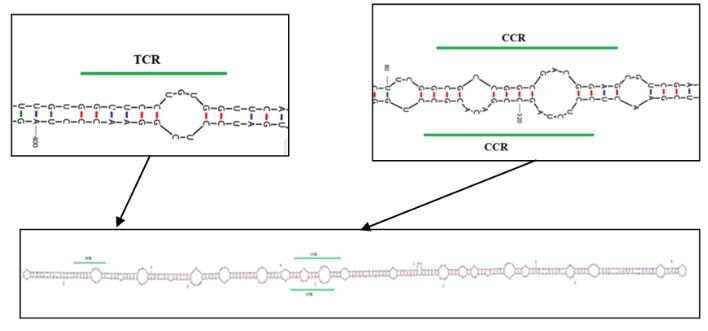
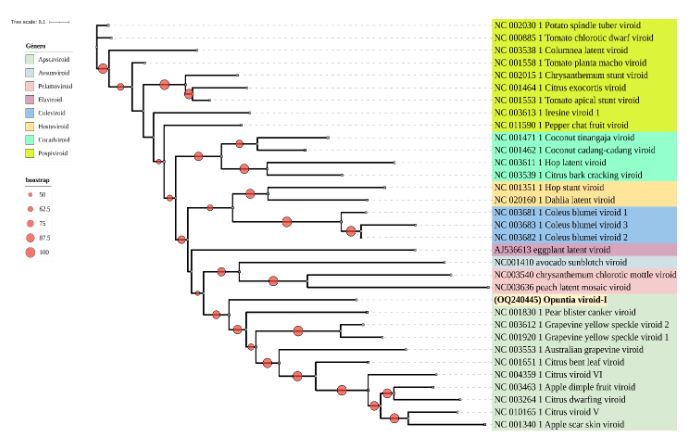
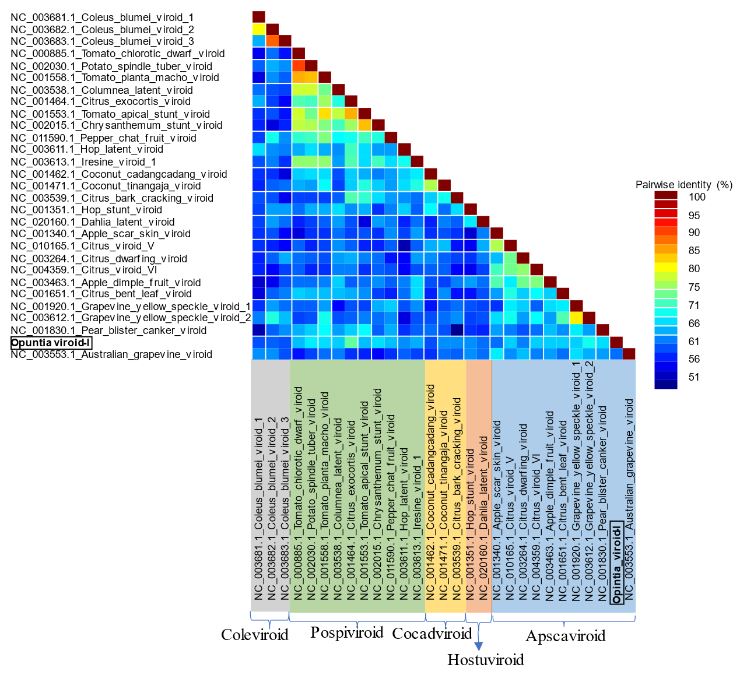
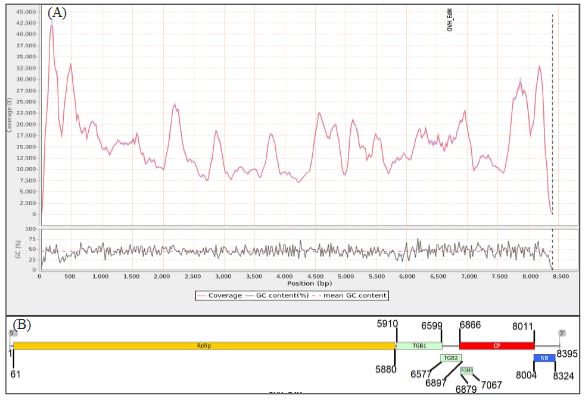
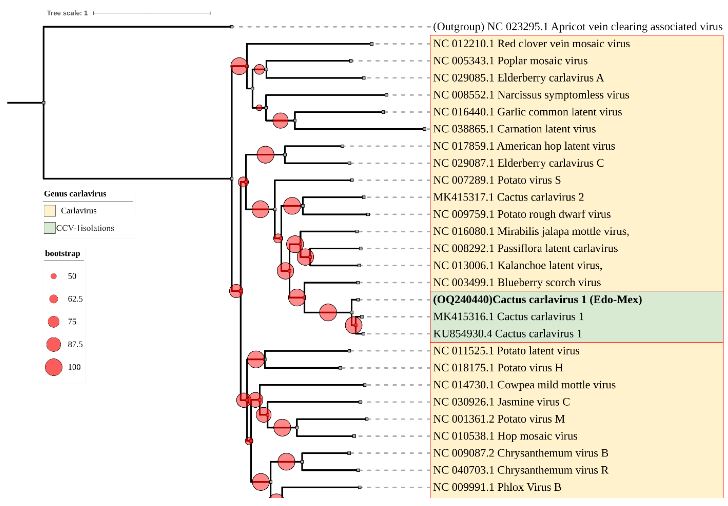
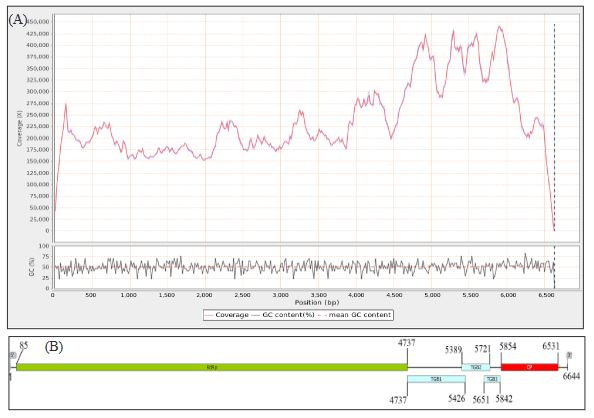
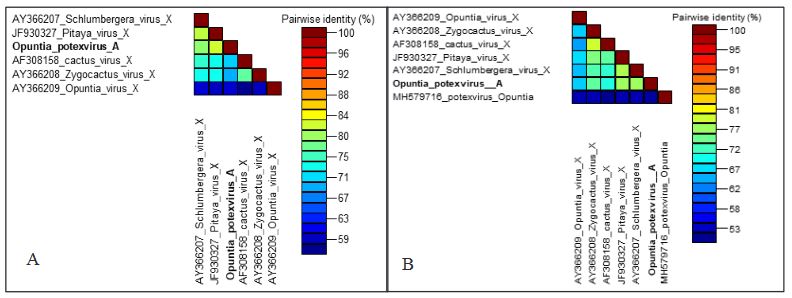
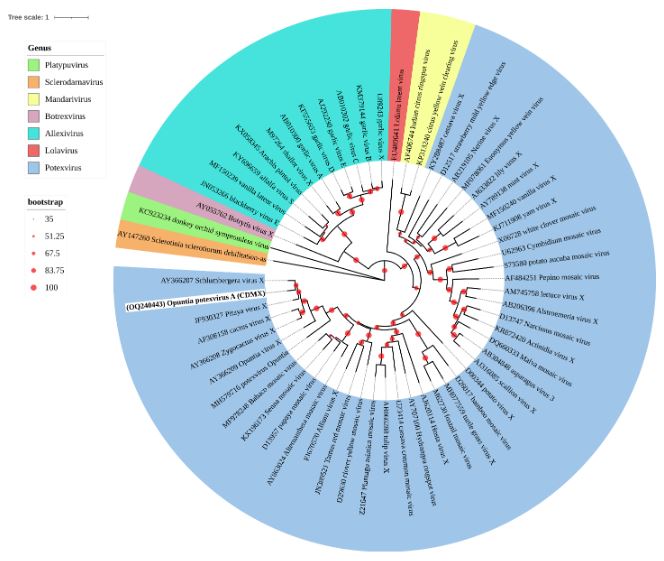
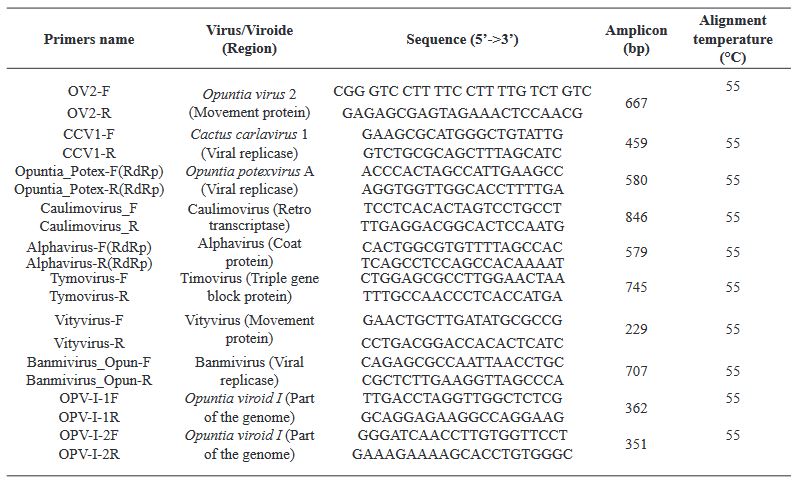
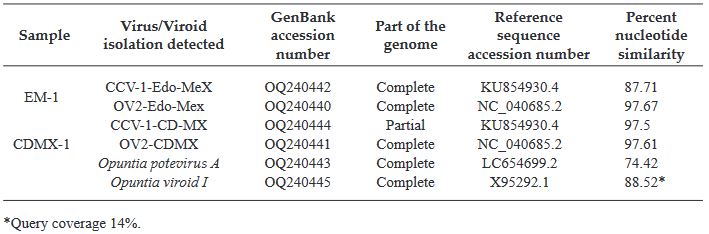
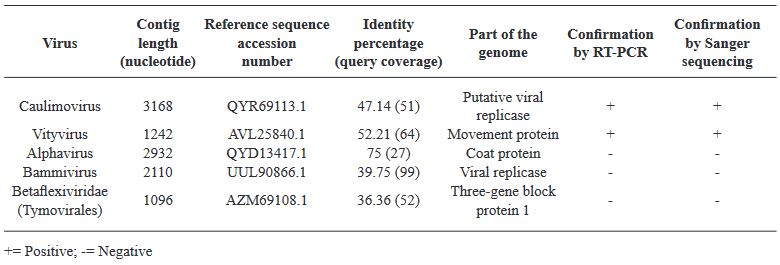
Results. In the sample from EDMX, the genomes of Opuntia virus 2 (OV2, genus Tobamovirus) and Cactus carlavirus 1 (CCV-1, genus Carlavirus) were detected and recovered. In the sample from CDMX, in addition to OV2 and CCV-1, a new viroid and potexvirus were detected. The former has a circular RNA genome with a length of 412 nt for which the name “Opuntia viroid I” (OVd-I) is proposed. The primary structure of this viroid showed a nucleotide sequence identity of less than 80% with any of the currently known viroids and a phylogenetic relationship with the genus Apscaviroid (Family Pospiviroidae) with which it shares conserved structural motifs.
Conclusion. The new potexvirus was named Opuntia potexvirus A (OPV-A), whose viral replicase sequence has a 77.7 % amino acid identity with Schlumbergera virus X. Finally, CCV-1 was detected in 93 (72 %) of 129 vegetable prickly pear cactus samples collected in the four entities.
Share this link via
Or copy link

Aspergillus oryzae: An opportunity for agriculture
byKaren Berenice García Conde, Ernesto Cerna Chávez, Yisa María Ochoa Fuentes*, Jazmín Janet Velázquez Guerrero
Received: 23/February/2023 – Published: 14/November/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2302-2
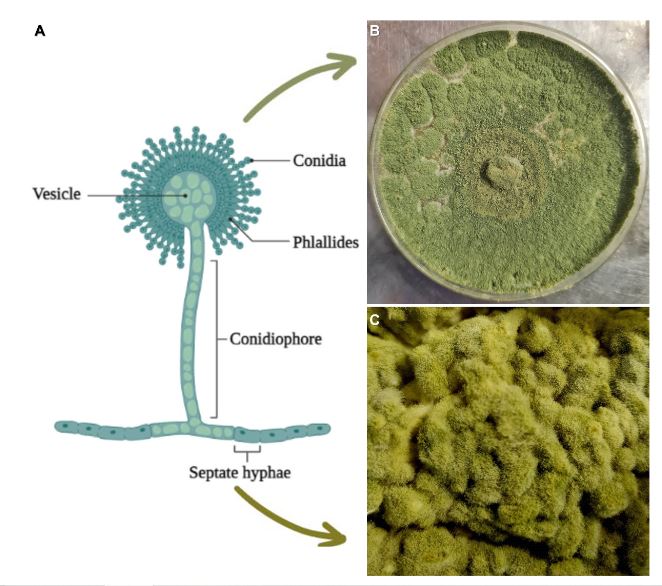
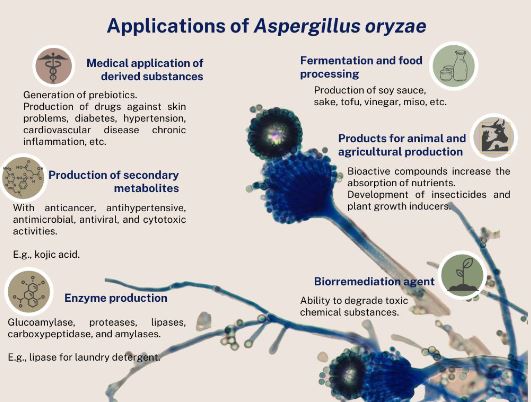
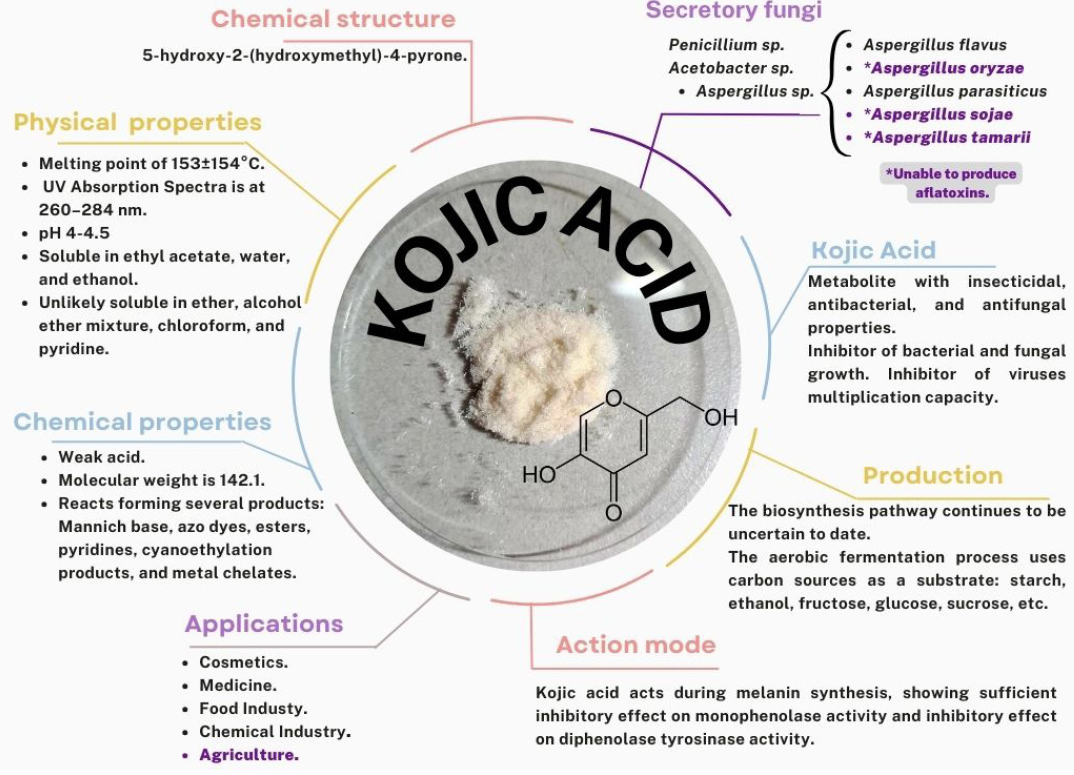
Abstract Aspergillus oryzae is a filamentous fungus capable of degrading various substances employing enzymes, which is why it is widely used in the biotechnological industry, pharmaceutical products, enzymes for industrial use, bleaching agents, anti-pollution textile treatments. However, few works focus on these microorganism’s field applications. This manuscript reviews the potentially beneficial applications of A. oryzae and some by-products in agriculture as biological control, growth inducer, and bioremediation for soils contaminated with heavy metals.
Share this link via
Or copy link
Etiology of rhizome rot of asparagus (Asparagus officinalis) in Atenco, Mexico State
byJuan Agustin Gonzalez Cruces*, José Sergio Sandoval Islas, Cristian Nava Díaz, Maricarmen Sandoval Sánchez
Received: 30/November/2023 – Published: 16/November/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2211-1
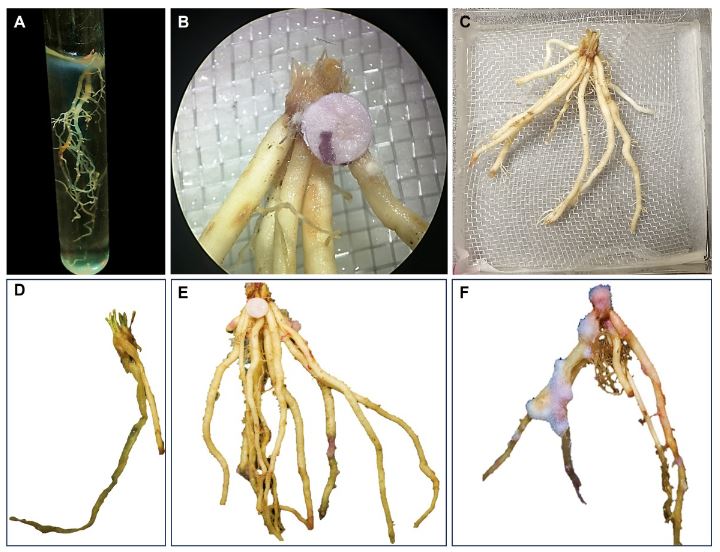
Abstract Background/Objective. The objective of this research was to identify the causal agent of asparagus rhizome rot, as well as evaluate different inoculation methods and the severity of the isolates.
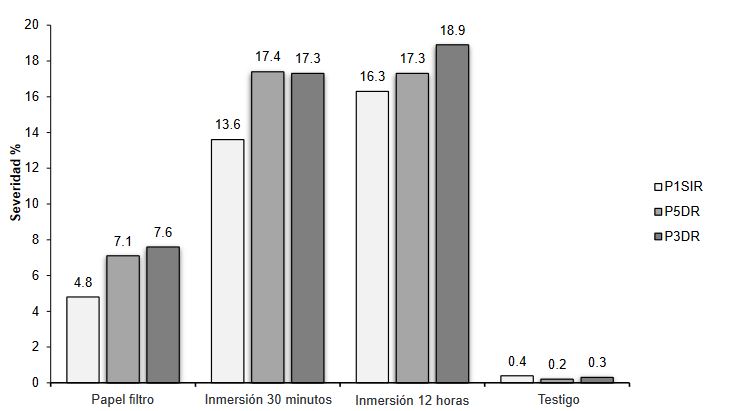
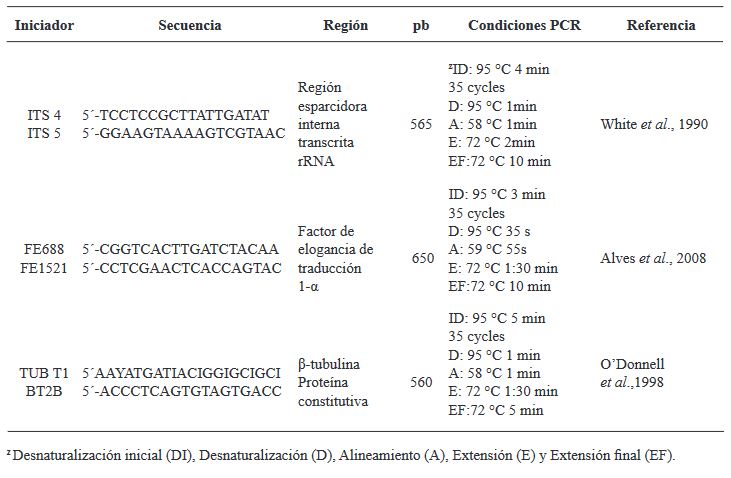
Materials and Methods. Sampling was carried out in five producing plots Atenco, Edo. from Mexico. Five isolates of Fusarium spp. were selected. (one per plot) to perform pathogenicity tests. Three isolates were selected for their colonization characteristics for severity tests with different inoculation methods: Immersion for 12 h, immersion for 30 min and inoculation by contact with absorbent paper soaked in 1 mL of inoculum. Concentrations of 1x106 conidia mL-1 were used. 10 rhizomes were used per treatment and 10 rhizomes without inoculation. To determine the severity, photographs (in GIMP®) of the rhizome were analyzed seven days after inoculation. The isolates were molecularly identified with ITS4/ ITS5, EF688/EF1521 and TUBT1/BT2B.
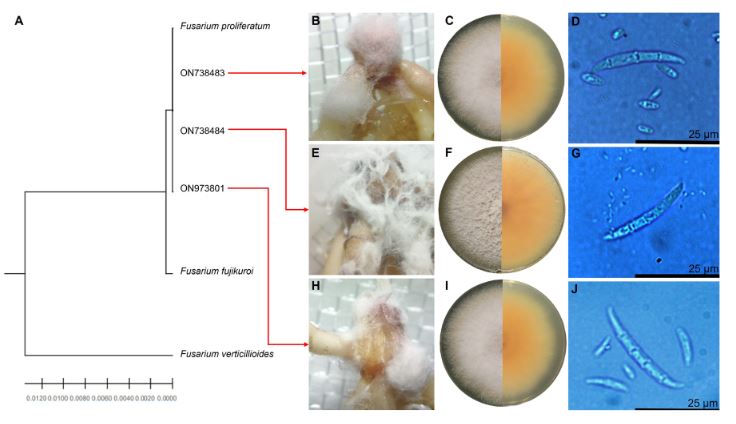
Results. Fusarium prolifetatum was morphologically and molecularly identified in the three isolates. The P3DR isolate was the most severe (14.6%), followed by P5DR (13.9%) and P1SIR (11.6%).
Conclusion. The most effective inoculation method was immersion for 30 min. They were registered in the NCBI Gene Bank with accessions ON738484 (P3DR), ON973801 (P5DR) and ON738483 (P1SIR). This is the first report of F. prolifetatum in the Edo. from Mexico.
Share this link via
Or copy link

Diagrammatic scale to evaluate the severity of gray mold (Botrytis cinerea) in pomegranate
byAlberto Patricio Hernández, Yuridia Mercado Flores*, Alejandro Téllez Jurado, María del Rocío Ramírez Vargas, Andrés Quezada Salinas
Received: 30/October/2022 – Published: 15/June/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2302-9
Abstract The aim of this study was to design and validate a diagrammatic scale to estimate the severity of gray mold induced by Botrytis cinerea in pomegranate cultivation. A total of 120 healthy and diseased fruits with varying degrees of affliction were collected from orchards with active production located in the municipalities of Chilcuahutla and Taxquillo in the state of Hidalgo, Mexico (20° 18’ 11’’ N, 99° 14’ 23’’ W, 20° 32’ 01’’ N, 99° 20’ 03’’ W, respectively). From these, 60 were selected to determine the severity percentage, according to a 6-class scale (Class 0 = 0%, Class 1 = >0% - 5% - 10%, Class 2 = >10% - 25% - 50%, Class 3 = >50% - 75% - 85%, Class 4 = >85% - 90% - 95%, and Class 5 = >95% - 100%), using the 2LOG software. With the obtained data, representative images were selected to build the diagrammatic scale using Adobe Photoshop. The accuracy (r2), precision (β0), and reproducibility (β1) were verified by simple linear regression applied to the data obtained by 12 evaluators with and without experience in the observation of plant diseases. As a result, values of r2 of 0.42 and 0.85 were obtained, without and with the use of the scale, respectively, which confirmed that this tool is suitable to evaluate the severity of the disease accurately and reproducibly.
Share this link via
Or copy link
There are no images, at the moment, loaded for this article.
In vitro antagonism of Trichoderma against Rhizoctonia solani
byJesús Orlando Pérez González, Sergio Gavino Ramírez Rojas, Ramiro Rocha Rodríguez, Katya Ornelas Ocampo, Jorge Miguel Vázquez Alvarado, Filogonio Jesús Hernández Guzmán, Mariel Garduño Audelo*
Received: 30/October/2022 – Published: 15/June/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2304-2
Abstract Trichoderma spp., is a highly efficient antagonist of root pathogens, such as Rhizoctonia solani, which causes loss in many crops. The aim of this research was to evaluate in vitro the antagonistic capacity of T. viride, T. koningii, T. harzianum and Trichoderma spp. isolates against R. solani from a potato crop. In confrontation tests, all Trichoderma isolates were classified as antagonists class 2 according to Bell scale, where T. harzianum and T. koningii showed more than 60% inhibition of the radial growth of R. solani at 120 h. In the interaction between T. harzianum and Trichoderma spp. with R. solani, as mycoparasitism strategy, vacuolization, lysis, coiling, and penetration were demonstrated, the last two were present in all Trichoderma isolates evaluated.
Share this link via
Or copy link
There are no images, at the moment, loaded for this article.
byOsvaldo Jhosimar Couoh Dzul, Karla Gisel Carreón Anguiano, Blondy Canto Canché*
Received: 30/October/2022 – Published: 15/June/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2210-5
Abstract The control of phytopathogens is key for food security. In the last decade, the use of interference RNA (iRNA) has been proposed as a technological tool for controlling diseases and pests in agriculture. Although different approaches have been described, such as the use of “Host-Induce Gene Silencing” (HIGS) and “Virus-Induce Gene Silencing” (VIGS), more recently a non-transgenic and environmentally friendly approach has emerged, called “Spray -Induce Gene Silencing” (SIGS), which uses double-stranded “naked” RNA (dsRNA). This review discusses recent reports on the use of dsRNA, especially SIGS, to control phytopathogenic fungi; emphasizing factors such as efficacy, safety in terms of human health and its stability in the environment. It also focuses on important phytosanitary problems in Mexico and Latin America that can be addressed with SIGS. This review concludes that SIGS technology has real potential to be used to control phytopathogenic fungi on plants in the field and on postharvest fruits. At the end, the critical tasks and the lines of research that must be carried out to promote the SIGS to make it a reality are considered.
Share this link via
Or copy link
There are no images, at the moment, loaded for this article.
byEdna Esquivel Miguel, José Luciano Morales García*, Martha Elena Pedraza Santos, Ana Tztzqui Chávez Bárcenas, Soledad García Morales, Samuel Pineda Guillermo
Received: 01/May/2023 – Published: 24/August/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2302-3
Abstract Elsinoe perseae (= Sphaceloma perseae) is the causal agent of the Mancha purpura or scab in avocado (Persea americana). In this study, the pathogenicity and virulence of E. perseae isolates from different agroecologicalproducing areas of Michoacán, Mexico will be reduced. For this, nursery plants with avocado fruits of Flor de María and Méndez varieties were used. On the other hand, the in vitro sensitivity of chemical fungicides (Azoxystrobin, thiabendazole, Pyraclostrobin, Cyprodinil + Fludioxonil and Azoxystrobin + Propiconazole) and authorized for use in orchards with organic management (copper sulfate, copper gluconate, copper oxychloride and the plant extract Larrea tridentata). The observed symptoms of Mancha purpura in the inoculated fruits were corroborated with those described for E. perseae in avocado. Inoculated fruits Flor de María variety shows the highest susceptibility to the pathogen. The isolates of E. perseae presented different degrees of virulence. The isolates showed different in vitro sensitivity values to the fungicides evaluated in the experiment. The pathogen showed the most sensitivity in vitro to chemical fungicides: thiabendazole and Azoxystrobin + Propiconazole (100% inhibition), and to those authorized in orchards with organic management: L. tridentata and copper oxychloride (on average 58% inhibition).
Share this link via
Or copy link
There are no images, at the moment, loaded for this article.
Effect of adjuvants, fungicides and insecticides on the growth of Trichoderma koningiopsis Th003
byYimmy Alexander Zapata Narváez*, Blanca Lucia Botina Azain
Received: 01/May/2023 – Published: 24/August/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2305-1
Abstract The effect of 44 agrochemicals (fungicides, insecticides and adjuvants) on the mycelial growth and germination conidia of Trichoderma koningiopsis Th003 was determined by seeding of 5 mm discs of fungal mycelium in Sabouraud agar supplemented with each agrochemical and seeding of conidia suspended in solutions of agrochemicals in water agar. For the adjuvants, their effect on the phyllospheric establishment of T. koningiopsis Th003 was determined by following their population in cape gooseberry leaflets inoculated with the fungus suspended in them. Eight fungicides did not inhibit the fungus mycelial growth or the conidia germination. Fenhexamid - Tebuconazole, Flutriafol and Kasugamicina inhibited it between 34 and 48% without affecting germination, Thiram - Pyrimethanil, Prochloraz, Tiabendazol, Spiroxamina and Triadimenol - Tebuconazole inhibited the growth and Thiram - Pyrimethanil and Dodine did not allow conidia germination. Insecticides and adjuvants presented an inhibition of up to 70% but did not affect the conidia germination. No negative effects of the adjuvants on the phyllosphere establishment of T. koningiopsis Th003 were observed, recovering from the treatments approximately 1x103 CFU g-1.
Share this link via
Or copy link
There are no images, at the moment, loaded for this article.