


Share this link via
Or copy link


Article filters
Search Papers
Estimation of losses caused by Potato virus Y in potato crop in Coahuila
byJoel De Santiago Meza*, Gustavo Alberto Frías Treviño, Luis Alberto Aguirre Uribe, Alberto Flores Olivas
Received: 05/April/2024 – Published: 30/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2404-2
Abstract Background/Objective. The objective was to experimentally evaluate the losses caused by PVY in the Fianna variety potato crop and, consequently, estimate the losses caused by this virus in the potato-producing area of Coahuila.
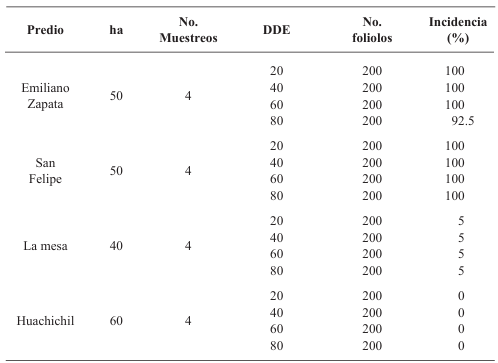
Materials and Methods. Furrows of an experimental plot planted with potato seedling and seed-tuber, were mechanically inoculated with PVY at 20, 40, 60 and 80 days after emergence. The tubers produced were harvested and losses in each treatment were evaluated. Additionally, in four commercial potato fields in this same state, leaflet samples were taken at 20, 40, 60 and 80 days after the emergence, and the percentage of plants infected with PVY was evaluated by ELISA tests. Loss data from the experimental plot and incidence data from the farms were used to develop a statistical model to estimate losses caused by PVY in the Coahuila region.
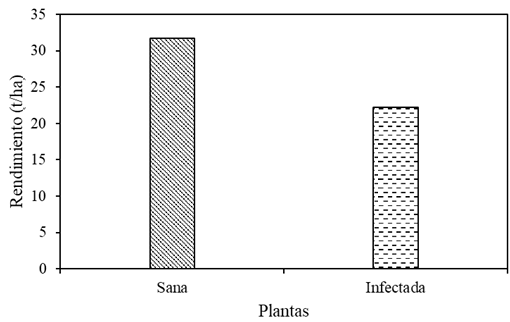
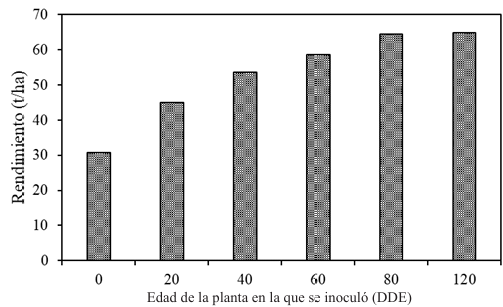
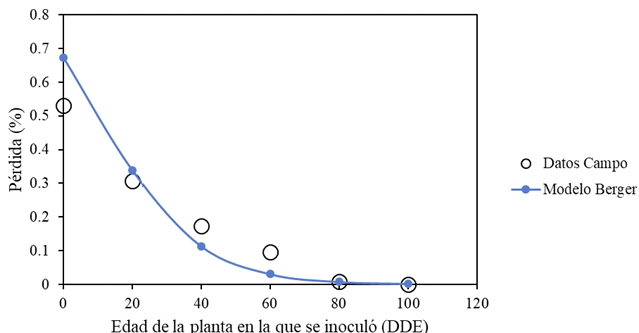
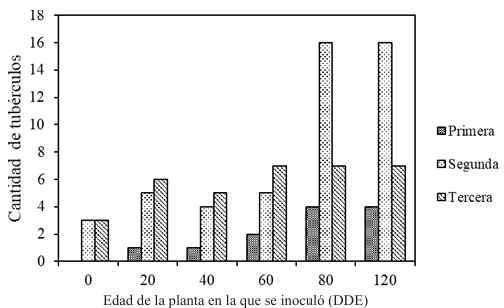
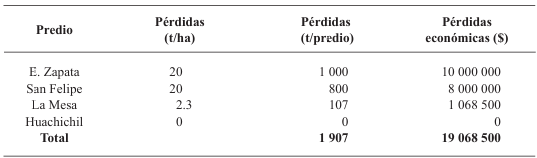
Results. Yield losses due to PVY in the experimental plot were 9.4% to 53%. The percentage of incidence of infected plants in commercial properties varied from 0% to 100%. The model that best fit the data obtained was Berger's Y=1/[1+e (-{ln[yo/(1-y0)]+r*dae})]. The estimated losses in the Coahuila region in the 2022 cycle were 18%, equivalent to $19 068500.
Conclusión: This information highlights the importance of using certified PVY-free seed and protecting the crop from emergence until 60 DAE.
Share this link via
Or copy link


Molecular aspects of phaseolotoxin biosynthesis produced by Pseudomonas syringae pv. phaseolicola
byAlejandra Chacón López, José Luis Hernández Flores, Efigenia Montalvo González, Selene Aguilera Aguirre*
Received: 20/August/2024 – Published: 30/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2308-2
Abstract Background/Objective. Phaseolotoxin is produced by one of the most important and studied phytopathogens in the agricultural area: Pseudomonas syringae pv. phaseolicola. This bacterium causes halo blight, a disease that devastates the bean crop. The success of P. syringae pv. phaseolicola is related to its genetic information, which allows it to synthesize deleterious metabolites for its host, such as phaseolotoxin. This research aimed to analyze the molecular basis of the mechanism of action, immunity, genetics involved in the biosynthesis of phaseolotoxin, molecular diagnostic strategies, and molecular techniques developed in Mexico to manage bean halo blight.
Materials and Methods. The search and analysis of the most relevant scientific information regarding the biosynthesis of phaseolotoxin and the molecular studies of the pathogenicity and virulence factors of P. syringae pv. phaseolicola has contributed to the development of molecular strategies focused on the diagnosis and management of halo blight in beans.
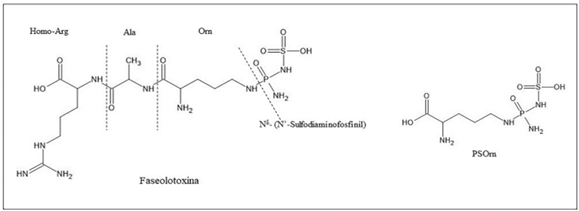
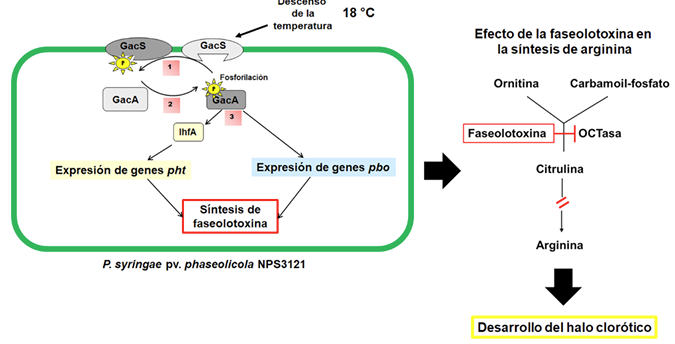
Results. P. syringae pv. phaseolicola produce phaseolotoxin, responsible for forming the chlorotic halo characteristic of halo blight, this toxin is an inhibitor of OCTase, an enzyme that participates in the arginine synthesis pathway in beans. The Pht and Pbo chromosomal regions contain genes involved in the synthesis and immunity of phaseolotoxin, and the expression of these genes is regulated by the GacS/GacA system and temperature. The identification of genes involved in the synthesis of pathogenicity and virulence factors, such as phaseolotoxin, has allowed the development of strategies for diagnosis and management of the disease based on DNA amplification and the use of molecular markers that facilitate the identification of bean cultivars resistant to the pathogen.
Conclusion. Molecular studies have contributed to understanding how the phaseolicola pathovar produces phaseolotoxin. This information has been essential to understanding how bacteria have evolved from non-pathogenic to pathogenic variants. In addition, they provide information that allows the development of new strategies for timely diagnosis and contributes to strategies for managing halo blight.
Share this link via
Or copy link




Toxicity of contact fungicides to four Trichoderma species: an in vitro compatibility approach
byConrado Parraguirre Lezama, Omar Romero Arenas*, Alba Cruz Coronel, Amparo Mauricio Gutiérrez, Carlos A Contreras Pare, Antonio Rivera Tapia
Received: 25/February/2024 – Published: 27/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2402-7
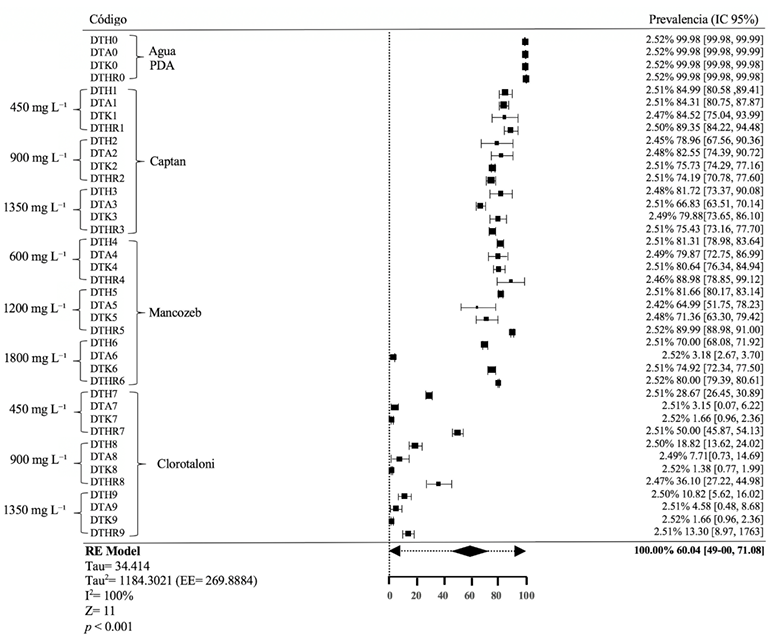
Abstract Objective/Background. The transition towards responsible agricultural practices is essential to promote the health of agroecosystems and ensure food security. Promoting comprehensive research that combines chemical and biological methods represents a significant advance in the management of phytopathogens. This novel approach is based on the premise that the joint action between fungicides and an antagonistic agent such as Trichoderma spp. can offer robust protection compared to individual approaches. The objective of the study is to investigate the in vitro resistance and compatibility of four Trichoderma species against three fungicides widely used in Mexico.
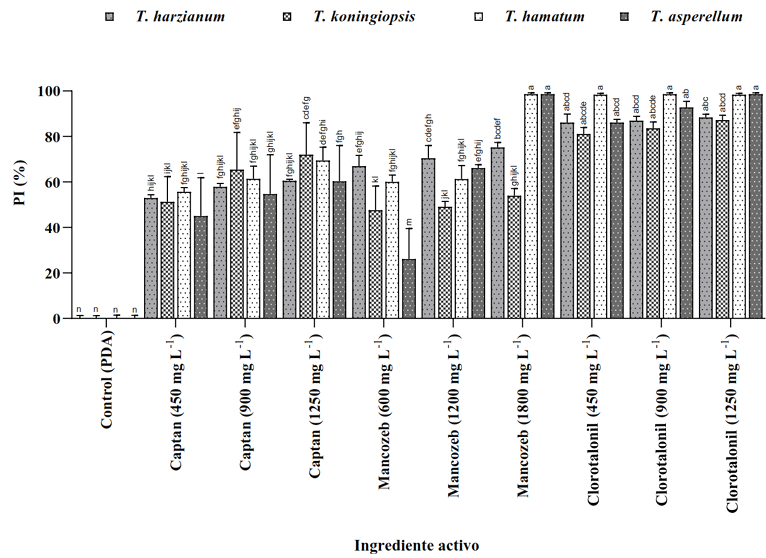
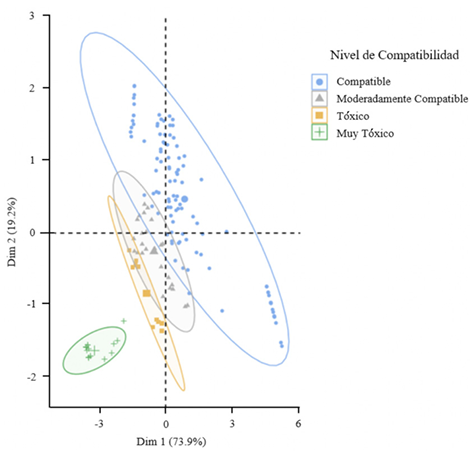
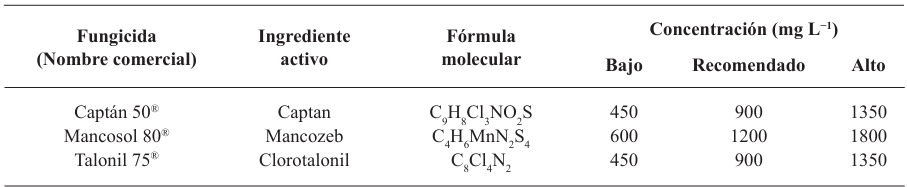
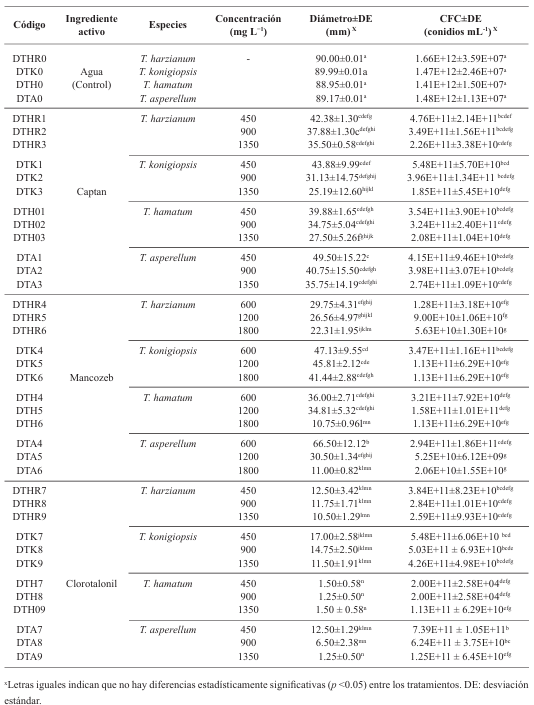
Materials and Methods. The controlled poisoning technique was used in PDA medium under controlled conditions with three concentrations (450, 900 and 1350 mg L−1) for the active ingredients Captan and Chlorothalonil, while for Mancozeb 600, 1200 and 1800 mg L−1 were used. Compatibility was determined in relation to the control group using the statistical software SPSS Statistics version 26 for the Windows operating environment.
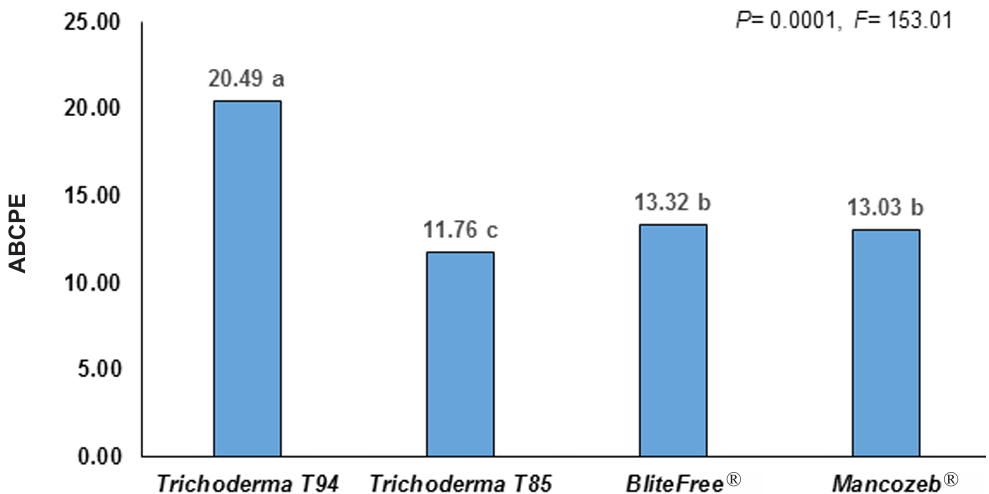
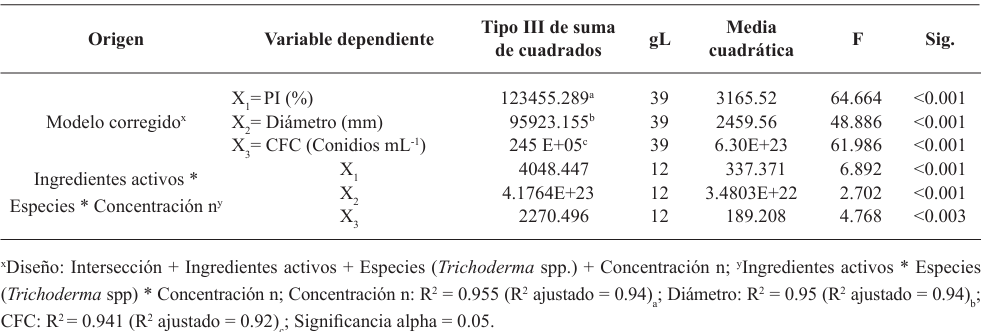
Results. The study revealed that the strains of T. harzianum, T. hamatum, T. koningiopsis and T. asperellum exhibited an overall compatibility of 60.04% for the active ingredients evaluated, with the fungicide Captan 50® showing the highest percentage of compatibility (79.87%) at concentrations of 450, 900 and 1350 mg L–1. T. harzianum showed greater tolerance to the active ingredient Chlorothalonil at a concentration of 450 mg L-1, however, at higher concentrations it showed greater toxicity, with T. koningiopsis exhibiting the lowest resistance at its three tested concentrations.
Conclusion. Treatments with different concentrations of the fungicides Captan, Mancozeb and Chlorothalonil showed a marked variability in terms of prevalence and toxicity towards the tested Trichoderma species in vitro. This approach allows the design of integrated management strategies minimizing the dependence on chemical products and promoting compatibility between biological agents and fungicides.
Share this link via
Or copy link
Etiology of brown rot in strawberry (Fragaria x nanassa) in the State of Mexico
byHugo Velasco Montaño, Victoria Ayala Escobar, Daniel Téliz Ortiz, Nadia Landero Valenzuela*, Santos Gerardo Leyva Mir
Received: 07/June/2024 – Published: 27/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2406-3
Abstract Background/Objective. In a strawberry crop established in a greenhouse in Montecillo, Texcoco, State of Mexico, in 2022, brownish brown leaf spots and rotting of fruits with asymmetric sunken lesions were observed, which extended and acquired a brown color. The objective of the present work was to identify the causal agent of brown rot in fruits and strawberry plants.
Materials and Methods. Symptomatic fruits and leaves were collected, from which fungal isolates were obtained to perform pathogenicity tests on plants and fruits, in plants by two inoculation methods: spraying via foliar and via root; in fruits by immersion. Concentrations of 2×106 conidia mL-1 were used. The ITS region of the rDNA was amplified and sequenced by PCR with the universal primers ITS1-ITS4.
Results. Pilidium concavum was morphologically and molecularly identified as the causal agent of brown spot and brown rot on strawberry. It was found to be pathogenic in strawberry fruits cv. Aromas and in plants less than two months old. It showed variation in virulence, in affected plants it varied from 40 to 50%, in fruits it reached 100%.
Conclusion. The result determines that Pilidium concavum is a pathogen that produces brown leaf spot and brown rot in strawberry fruits. It allows new lines of research related to the impact of the disease on strawberry production, yield and quality in Mexico. This research is the first report of Pilidium concavum as a strawberry pathogen in the State of Mexico.
Share this link via
Or copy link


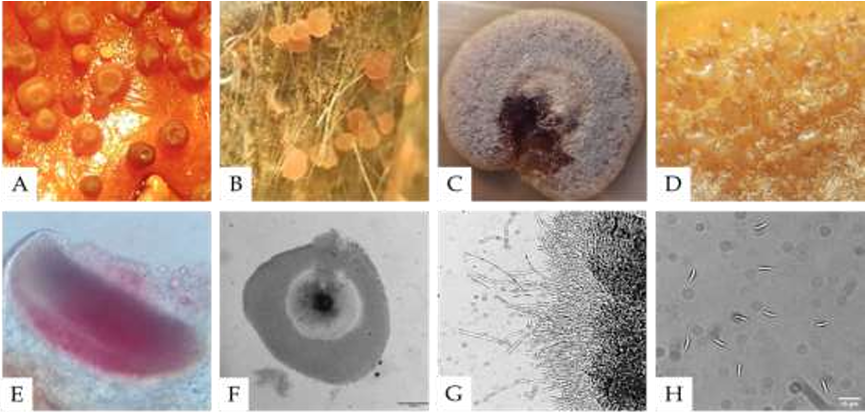




byValeria Valenzuela Ruiz, Fannie Isela Parra Cota, Gustavo Santoyo, María Isabel Estrada Alvarado, Luis Alberto Cira Chávez, Ernestina Castro Longoria, Sergio de los Santos Villalobos*
Received: 01/July/2024 – Published: 18/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-18
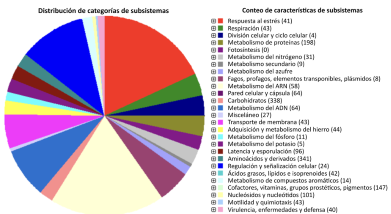
Abstract Background/Objetive. Bacillus paralicheniformis TRQ65 was isolated from wheat (Triticum turgidum subsp. durum) rhizosphere in commercial fields in the Yaqui Valley, Mexico. This strain was one of the most abundant bacteria in the rhizosphere. The objective of this study is to explore the potential biological control action mechanisms of Bacillus paralicheniformis TRQ65 against phytopathogenic fungi of agricultural importance, through genome sequencing and mining.
Materials and methods. The biocontrol activity of this strain was quantified through in vitro dual assays evaluating inhibition zones against 11 agronomically important fungi. A whole-genome analysis was conducted as genomic mining to evaluate its potential for biological control.
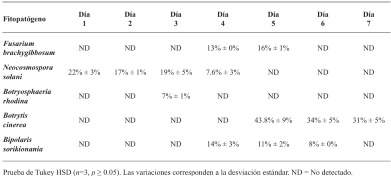
Results. Strain TRQ65 showed biocontrol activity against 45% of the studied fungi, where the highest inhibition was against Botrytis cinerea, 43.8% ± 9% on day 5. Based on genome sequencing and mining (antiSMASH), this bioactivity could be associated with the biosynthesis of lichenysin, bacillibactin, and/or fengycin.
Conclusion. This research provides the first insight into the potential biological control activity of strain TRQ65. Further studies need to be carried out to validate Bacillus paralicheniformis TRQ65 as an active ingredient in sustainable bacterial inoculants for eco-friendly agriculture.
Share this link via
Or copy link
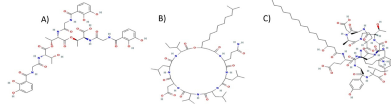
Mitigating Chili Plant Wilt: Synergy of Arbuscular Mycorrhizal Fungi and Silver Nanoparticles
byHilda Karina Sáenz Hidalgo, Esteban Sánchez Chávez, Nuvia Orduño Cruz, Mahendra Rai, Víctor Olalde Portugal, Graciela Dolores Ávila Quezada*
Received: 17/June/2024 – Published: 13/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-8
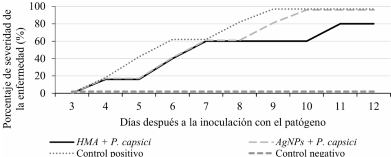
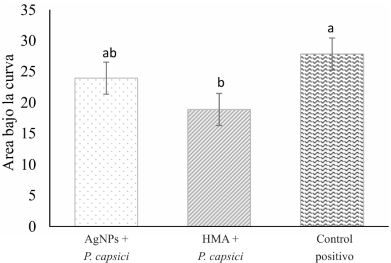
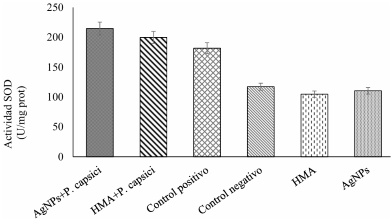
Abstract Background/Objective. The chilaca pepper (Capsicum annuum) is significantly impacted by the attack of the oomycete Phytophthora capsici, which causes chili wilt. Current methods for controlling this disease have been inefficient. Therefore, the search for more environmentally friendly alternatives is of great importance. In pursuit of this objective, we assessed the potential of arbuscular mycorrhizal fungi (AMF) and silver nanoparticles (AgNPs) to try to reduce or postpone chili pepper wilt.
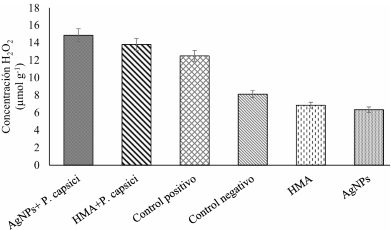
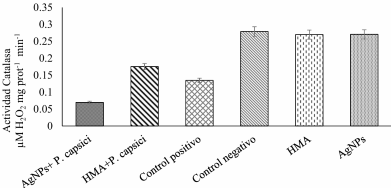
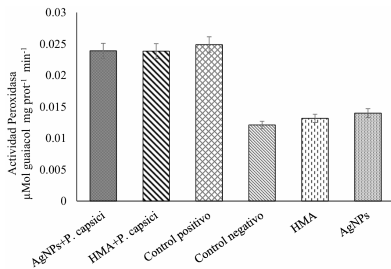
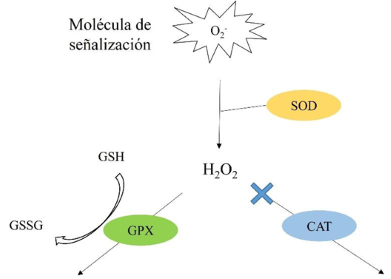
Materials and Methods.Growth parameters were measured in inoculated and non-inoculated chili pepper plants with AMF from a commercil consortium TM-73 (Biotecnología Microbiana) and the protective effects of AMF and AgNPs (NanoID®) against P. capsici as evaluated using a severity scale for wilt symptoms. Plant response to pathogen infection was assessed by measuring the activities of antioxidant enzymes: PER, SOD, CAT and H2O2.
Results. The results indicated that AMF application improved the growth parameters of C. annuum, while the plant-pathogen interaction induced an antioxidant enzymatic response. AMF maintained wilt symptoms at or below 80%, preventing plant death. Meanwhile, AgNPs (50ppm) delayed plant mortality compared to the control treatment.
Conclusion. The combined use of AMF and AgNPs offer options for future research in the disease management for chili peppers.
Share this link via
Or copy link


byFrancisco Palemón Alberto, Santo Ángel Ortega Acosta*, Erubiel Toledo Hernández, Cesár Sotelo Leyva, Guadalupe Reyes García, Elizabeth Tecomulapa Acatitlán
Received: 07/July/2024 – Published: 04/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-21
Abstract Background/Objective. Diseases of agricultural crops affect the yields and quality of products. Synthetic chemical compounds are generally used to control them; these cause harmful impacts to the environment, as well as to human health. In this sense, beneficial microorganisms can be used in agriculture as biocontrol agents, and contribute to obtaining food in sufficient and safe quantities. The fungus Irpex lacteus has been reported as a potential biocontrol agent. The objective of this research work was to evaluate the in vitro mycoparasitic capacity of the endophytic fungus I. lacteus P7B against 22 fungi and one oomycete associated with plant diseases.
Materials and Methods. The P7B isolate, previously detected as a mycoparasite, was used and molecularly identified by amplification and sequencing of the internal transcribed spacer (ITS) region of ribosomal DNA, using primers ITS1/ ITS4.The confrontations of the mycoparasite (P7B) against the phytopathogenic microorganisms were carried out in PDA culture medium. Three replicates were used for each microorganism, in addition to the controls, which consisted of placing the microorganisms individually.
Results. Molecular analyses determined that isolate P7B corresponded to Irpex lacteus (GenBank: PP922180). The results of the in vitro assays indicated that I. lacteus P7B inhibited all the phytopathogenic agents with which it was confronted, 100% inhibition by I. lacteus occurred approximately in 14 days, except for Rhizopus spp., this was at 23 days after the confrontations.
Conclusion. The present study demonstrates that the fungus I. lacteus presented 100% in vitro mycoparasitic capacity against the various fungi and an oomycete evaluated, so future work could focus on evaluating its mycoparasitic activity under field conditions.
Share this link via
Or copy link

byEstefanía Fonseca Chávez, Irvin Alonso Molina Marañón, Luz Irela Lugo Zambrano, Juan Carlos Martínez Álvarez, Guadalupe Arlene Mora Romero, Jesús Damián Cordero Ramírez, Karla Yeriana Leyva Madriga*
Received: 01/July/2024 – Published: 04/December/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-14
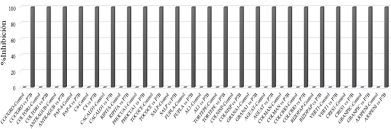
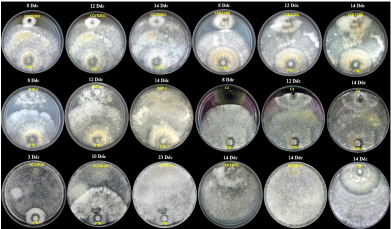
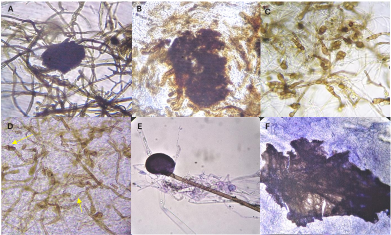
Abstract Background/Objective. Biological control offers new strategies for disease management in agriculture. In the present study, the antifungal activity of volatile organic compounds (VOCs) emitted by rhizobacteria, was evaluated in the growth and infective capacity of four isolates of E. turcicum obtained from symptomatic corn plants, in northern Sinaloa.
Materials y methods. The fungal isolates were characterized morphologically and molecularly, and their pathogenicity was corroborated in a detached leaf assay. The ability of VOCs to inhibit mycelial growth and infection of maize leaves by E. turcicum was evaluated in in vitro assays, in divided Petri dishes. Bacterial hydrogen cyanide production was qualitatively determined.
Results. The mycelial growth of E. turcicum was reduced by VOCs of at least one rhizobacteria, registering inhibitions between 22% and 63%. Leaves infection was reduced between 63% and 98% in the presence of rhizobacterial VOCs. Hydrogen cyanide production was detected in strains B3 and B9.
Conclusion. Strain B95 was more effective in reducing mycelial growth and infection by E. turcicum. The production of hydrogen cyanide could be involved in its antagonistic effect. In-plant tests are required to corroborate its effectiveness, as well as characterize its volatile profile.
Share this link via
Or copy link
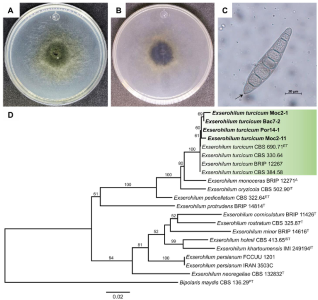
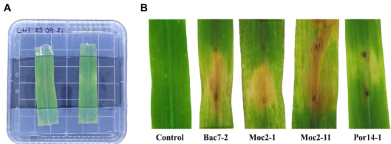


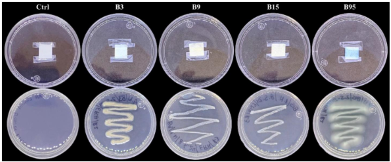
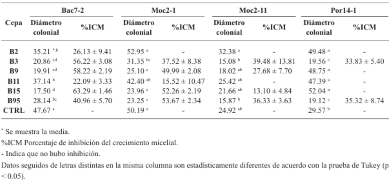
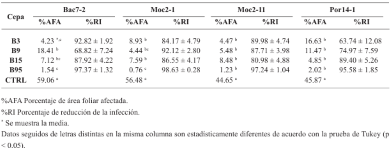
byJesús Eduardo Ramírez Méndez, Francisco Daniel Hernández Castillo, Gabriel Gallegos Morales, Diana Jasso Cantú, Roberto Arredondo Valdés, Marco Antonio Tucuch Pérez*
Received: 02/July/2024 – Published: 23/November/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-23
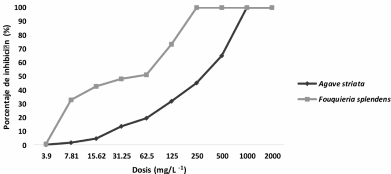
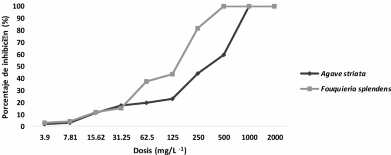
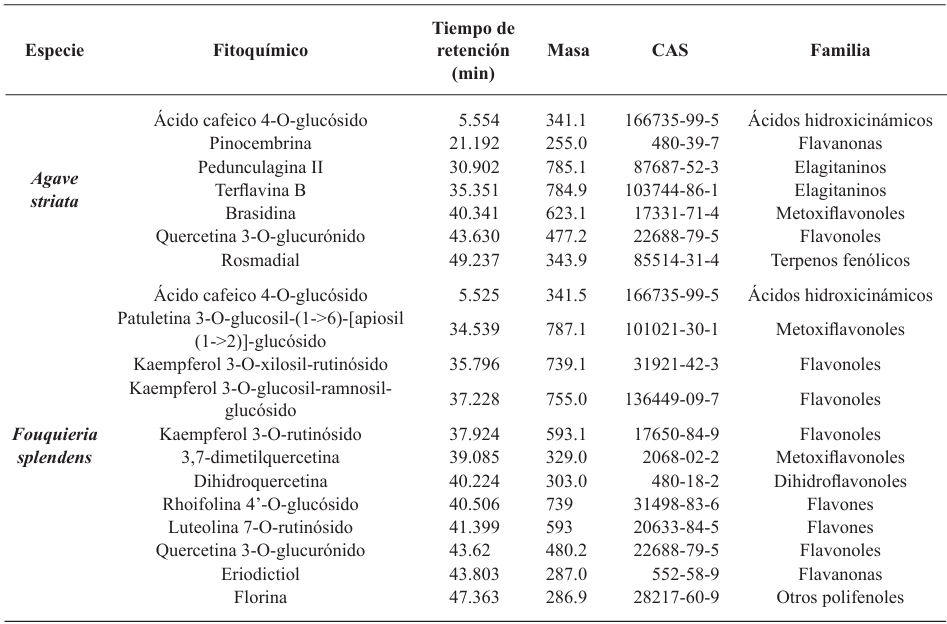
Abstract Background/Objective. Pythium aphanidermatum and Rhizoctonia solani are pathogens that affect agricultural crops. The objective of this study was to evaluate some Agave striata and Fouquieria splendens methanolic extracts, from the Chihuahuan Desert, against those fungi, in search for biological control alternatives.
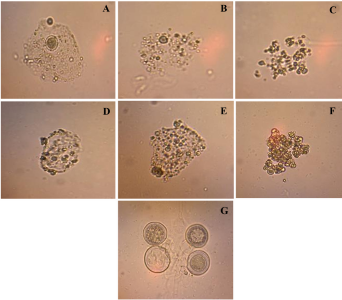
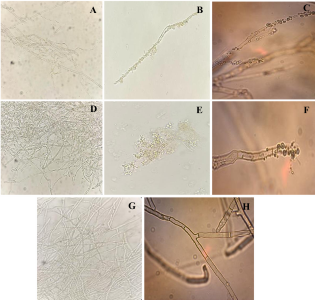
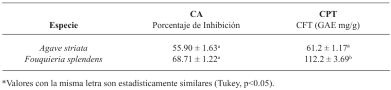
Materials and Methods. Pathogens from chili plants were isolated and identified using morphological and molecular methods. Methanolic extracts from both plants were prepared, and the antioxidant capacity (AC), the total polyphenol content (TPC), and the antifungal compounds via HPLC-MS were assessed. The antifungal effectiveness was tested at concentrations of 3.9-2000 mg L-1 using a poisoned medium assay, where fungi structural damage was observed.
Results. The F. splendens and A. striata extracts exhibited 55% and 68% AC, as well as 61 mg/g and 112 mg/g TPC, respectively. Both extracts contained caffeic acid and quercetin, while F. splendens also exhibited eriodyctiol, kaempferol, and luteolin; A. striata contained pinocembrin and theaflavin B. F. splendens attained 100% inhibition of P. aphanidermatum at 250 mg L-1, and of R. solani at 500 mg L-1, whereas A. striata achieved 100% inhibition at 1000 mg L-1 in both cases. The extracts produced lysis in P. aphanidermatum oogonia and mycelial fragmentation in R. solani.
Conclusion. The F. splendens and A. striata methanolic extracts demonstrate promising antifungal activity against P. aphanidermatum and R. solani, suggesting that these natural compounds might be useful as a biological alternative for pathogen control in agricultural crops.
Share this link via
Or copy link

byFannie Isela Parra Cota, Isabeli Bruno, Mónica García Montelongo, Sebastián González Villarreal, María Fernanda Villarreal Delgado, Liliana Carolina Córdova Albores, Alina Escalante Beltrán, Sergio de los Santos Villalobos*
Received: 10/July/2024 – Published: 19/November/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2024-34
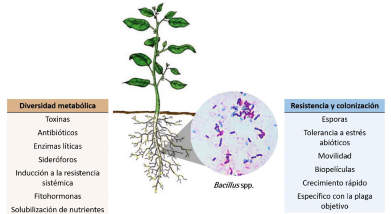
Abstract Background/Objective. Bacteria of the Bacillus genus have been studied since their discovery in 1872, for their ability to synthesize metabolites of interest such as proteins used to control phytopathogens. The objective of this research was to analyze the most outstanding species of the Bacillus genus, as well as the main compounds produced by these species, and the perspectives on the use of this bacterial genus for pest and disease control.
Materials and Methods. An exhaustive search was carried out in scientific articles and books to gather the most relevant information regarding the Bacillus genus, focusing on its role as a biological control agent for pests and pathogens.
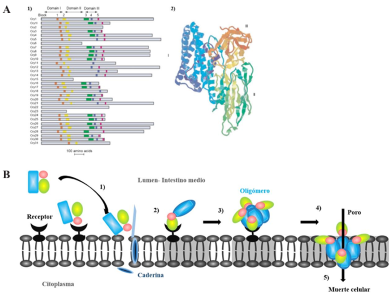
Results. The Bacillus genus includes more than 427 taxa, which can be classified into different groups. Among the biological control agents (BCA) are the Bacillus cereus group, which includes B. cereus, B. anthracis, and B. thuringiensis, and the B. subtilis group, which includes B. subtilis, B. licheniformis, and B. pumilus, mainly. B. thuringiensis, through cry genes, has molecular mechanisms to synthesize a crystalline inclusion during sporulation, which contains proteins known as endotoxins or Cry proteins. B. subtilis produces substances with a high potential for biological control, such as volatile organic compounds, as well as bioactive secondary metabolites.
Conclusion. The potential of the Bacillus genus to be used as biological control agents is evident. They are widely used for the development of different biopesticides that have advantages over other products. However, it is necessary to continue conducting research from the in vitro area in the laboratory to the field, to help guarantee their biosecurity and effectiveness.
Share this link via
Or copy link