
Share this link via
Or copy link


Article filters
Search Papers
byJosé Jesús Márquez Diego, Andrea Denisse Martinez Vidales, Errikka Patricia Cervantes Enríquez, Abraham Ruiz Castrejón, José Humberto Romero Silva, Maria Edith Ortega Urquieta, Fannie Isela Parra Cota, Sergio de los Santos Villalobos*
Received: 18/July/2023 – Published: 06/March/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2402-9
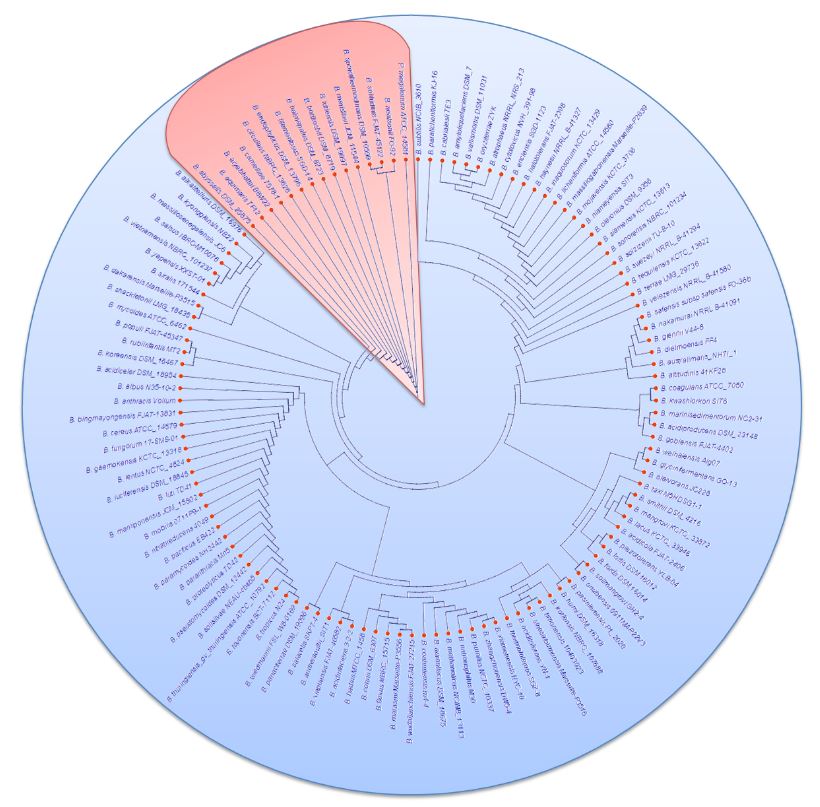
Abstract Background/Objective. Bacillus is a cosmopolitan bacterial genus with a great genome diversity. Thus, by exploring its genome background, it is possible to understand more about the physiological and biochemical traits involved in its biological control against phytopathogens. The objective of this work was to correlate the phylogenomic relationships of the type species of the genus Bacillus with the presence of gene clusters associated with biological control of plant pathogens, through genome mining.
Materials and Methods. Based on the literature, 336 species belonging to the genus Bacillus have been reported; however, after re-classification, a total of 123 type species have been recognized, and curated genomes were found in the EzBioCloud platform (http://www.ezbiocloud.net/). The overall genome relatedness indices (OGRIs) were used for this work, which indicate how similar two sequences of a genome are. Then, the Realphy platform was used to create the phylogenomic tree 1.13 (Action-based phylogeny constructor reference). Finally, the prediction of biosynthetic gene clusters (BGC) associated with the biological control of phytopathogens was carried out using antiSMASH v6.0 (https://antismash. secondarymetabolites.org/).
Results. The present strategy allowed us to correlate and predict the biological control capacity of the Bacillus species under study based on their taxonomic affiliation since at a shorter evolutionary distance from Bacillus subtilis a high potential capacity to produce biological control compounds was observed. However, the possibility that they acquire the ability to produce new biocontrol compounds during their evolutionary separation is not ruled out.
Conclusion. This work validates the correlation between the taxonomic affiliation of the studied Bacillus species and their biological control capacity, which is useful in the bioprospecting stage to design promising biopesticides.
Share this link via
Or copy link

byAlfonso Muñoz Alcalá, Gerardo Acevedo Sánchez, Diana Gutiérrez Esquivel, Oscar Bibiano Nava, Ivonne García González, Norma Ávila Alistac, María José Armenta Cárdenas, María del Carmen Zúñiga Romano, Juan José Coria Contreras, Serafín Cruz Contreras, Gustavo Mora Aguilera*
Received: 18/July/2023 – Published: 06/March/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2312-1
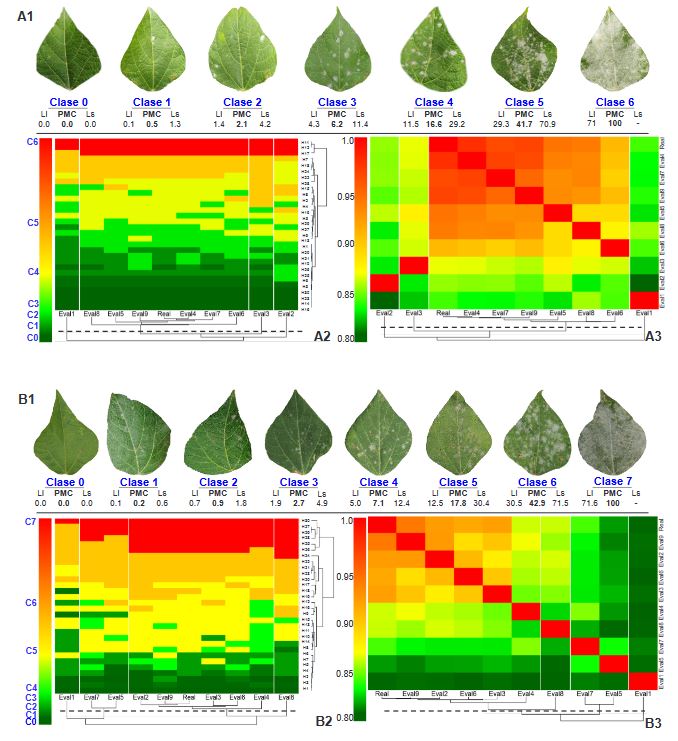
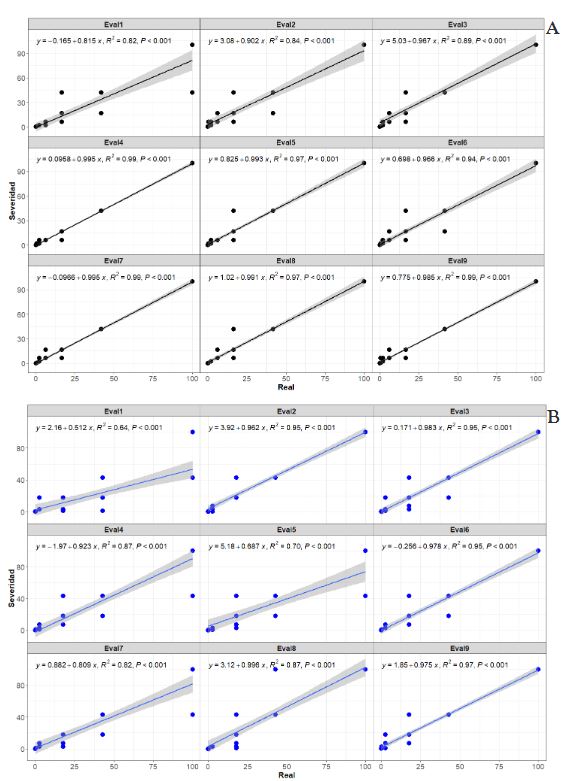
Abstract Background/Objective. Epidemiological research on Phaseolus coccineus is lacking. The aim was to develop and validate digital methods to quantify the severity associated with powdery mildew in ayocote bean.
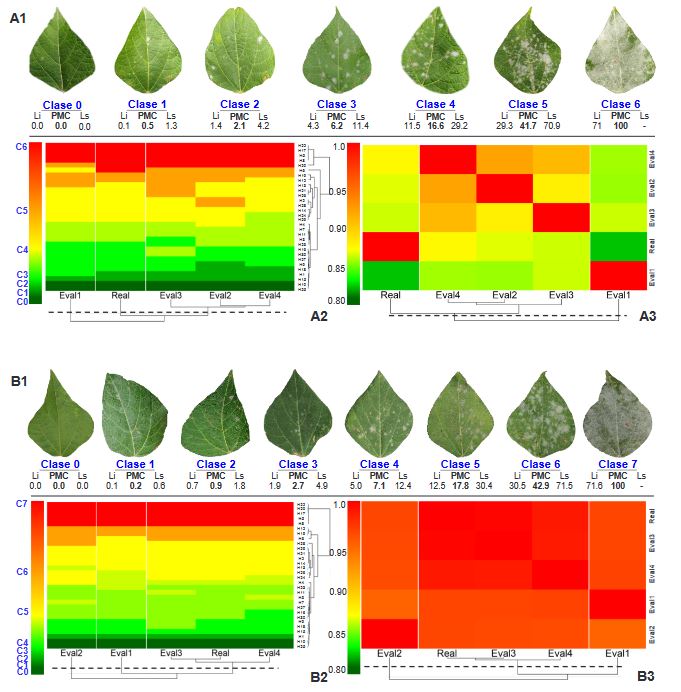
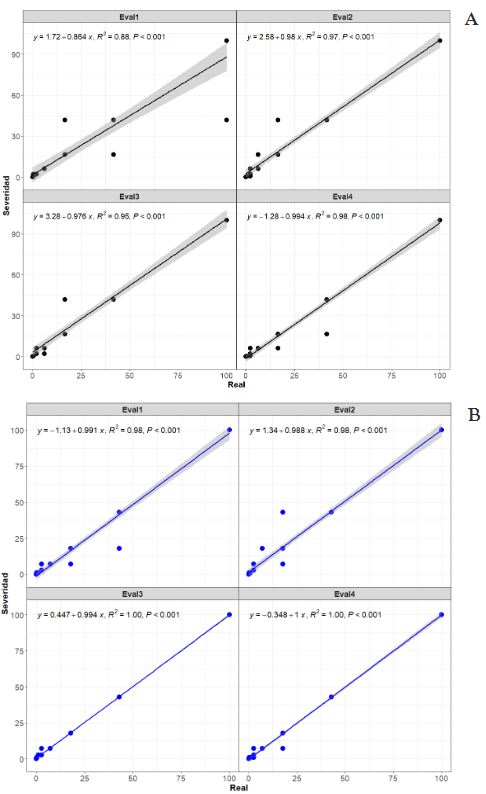
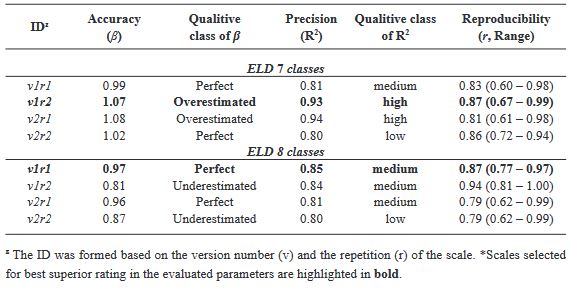
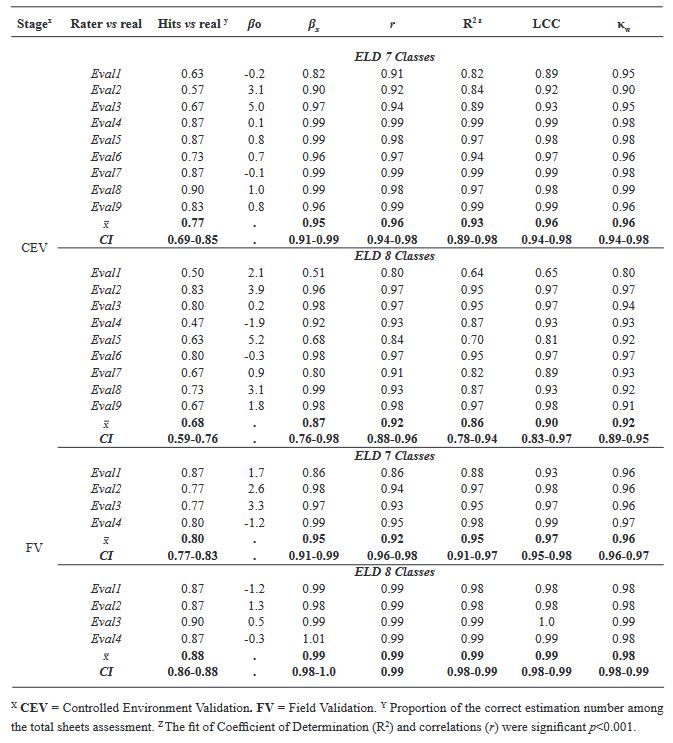
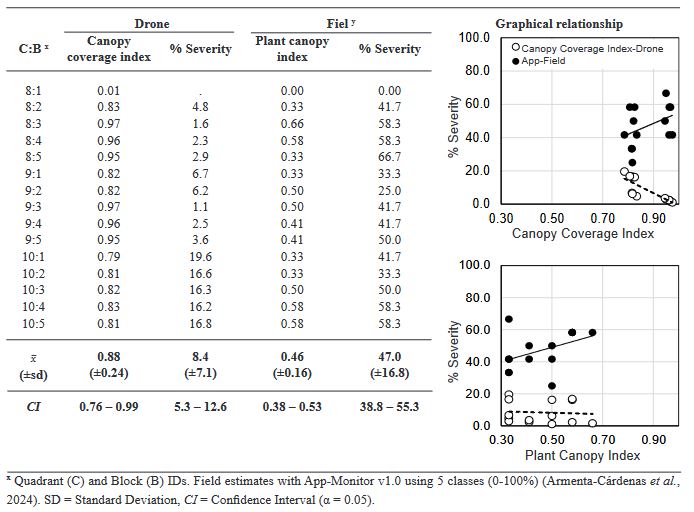
Materials and Methods. An ayocote bean plot with 65.3 % incidence and 22.7 % average powdery mildew foliar severity was selected. Based on 250 leaves collected in field with varying severity degrees, eight 7- and 8-class logarithmic-diagrammatic scales (ELD) were designed and validated in a controlled environment (CEV) and field (FV). In Rstudio®, accuracy (β), precision (R2), reproducibility (r), and agreement level were determined with Cohen’s kappa index (κw) and Lin’s concordance coefficient (LCC). Additionally, a Hierarchical Cluster Analysis (HCA) was performed by scale and assessment environment for clustering by similarity evaluation. In ArcMap® v10.3, in a 15-quadrant block, an ‘image segmentation’ analysis was performed using supervised classification and maximum likelihood to estimate powdery mildew severity and an indicator of canopy coverage index (VCI).
Results. In VEC-1, v1r2 (ELD-7c; β=1.07, R2=0.93, r=0.87) and v1r1 (ELD-8c; β=0.97, R2=0.85, r=0.87) scales were best evaluated. In VEC-2, comparing clusters conformed in the HCA, the ELD-7c was the best scored with perfect accuracy (β>0.96), very high precision (R2>0.94), very high reproducibility (r=0.97-0.99) and very high agreement (κw>0.96; LCC>0.97); and in ELD-8c reproducibility and agreement decreased. In VCa, ELD-7c maintained optimal metrics, but ELD-8c reached ideal parameters for preventive ELD in early stages of powdery mildew (β>0.98, R2>0.98, r=0.99, κw=0.99-0.999, LCC=0.98-0.999). Image analysis estimated severity = 8.4 % (CI = 5.3 - 12.6 %) and ICV = 0.88 (CI = 0.76 - 0.99), contrasting with field assessment 47 % (CI = 38.8 - 55.3 %) and 0.46 (CI = 0.76 - 0.99), respectively, mainly with ICV > 0.94 due to less symptomatic leaf exposure. Suggests applicability for canopy estimation with restrictions for severity based on pathogen expression.
Conclusion. A methodology for ELD development is proposed, comprising: image acquisition, processing and quantification; controlled validation and field validation. Validation statistics included precision (R2); accuracy (β); reproducibility (Pearson’s coefficient and Hierarchical Cluster Analysis); and agreement (Lin’s Coefficient and Kappa Index), proposed in a comprehensive approach for first time. RGB-drone images are proposed to estimate a comprehensive vigor and severity coverage index.
Share this link via
Or copy link


byNorma Ávila Alistac, Gustavo Mora Aguilera, Héctor Lozoya Saldaña*, Erika J. Zamora Macorra, Camilo Hernández Juárez
Received: 18/July/2023 – Published: 06/March/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2311-2
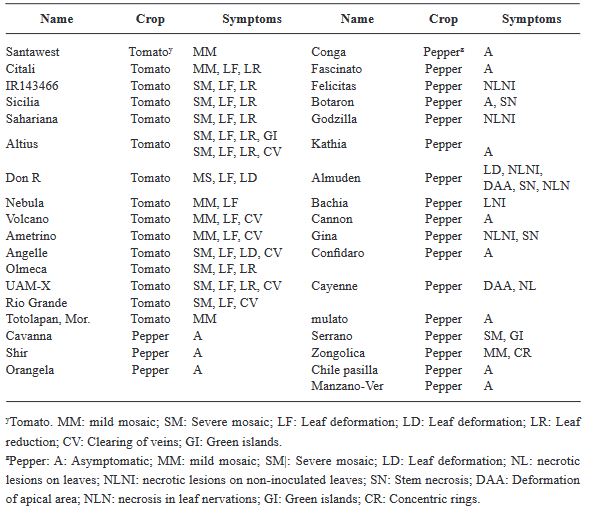
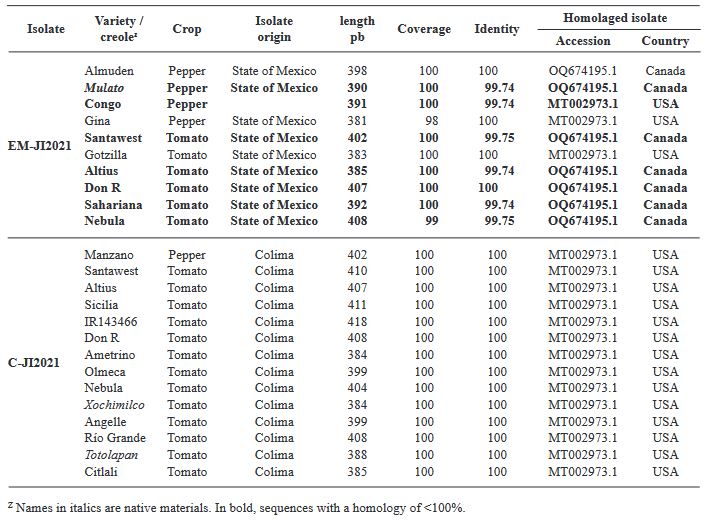
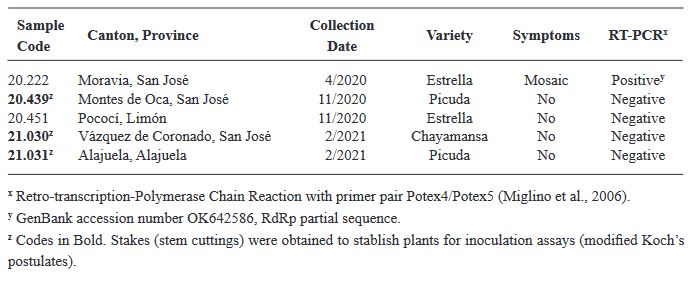
Abstract Background/Objective. The objective was to analyze the variability of two Mexican isolates of ToBRFV after a process of inoculation and multiplication in different commercial and Mexican landraces of tomato (Solanum lycopersicum) (15 materials) and pepper (Capsicum annuum) (20 materials), and to evaluate the expression of symptoms under greenhouse conditions.
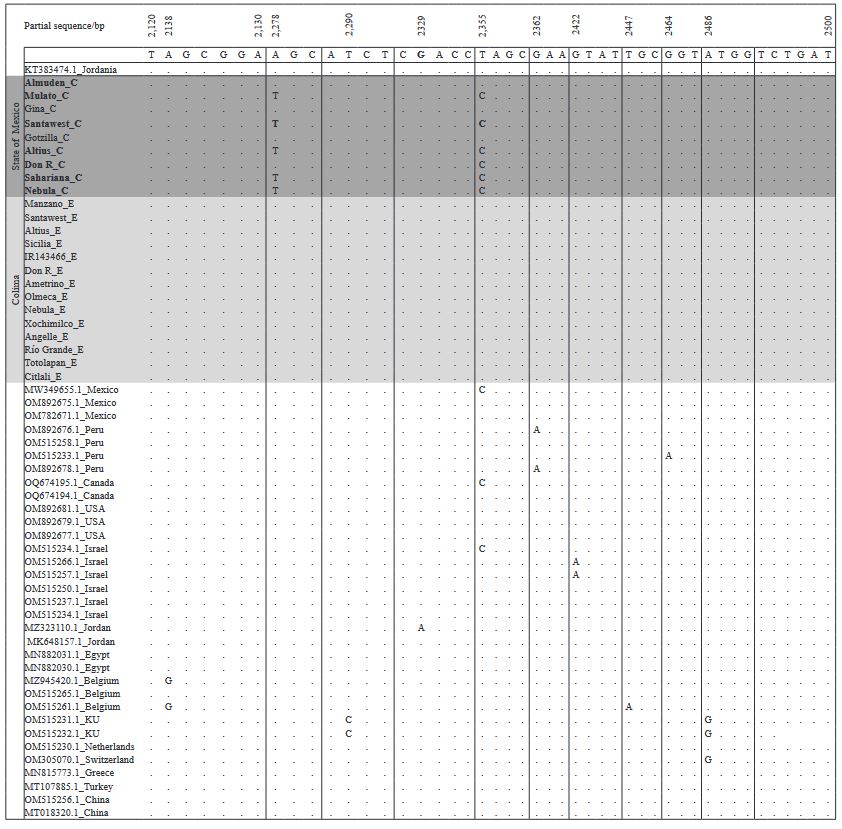
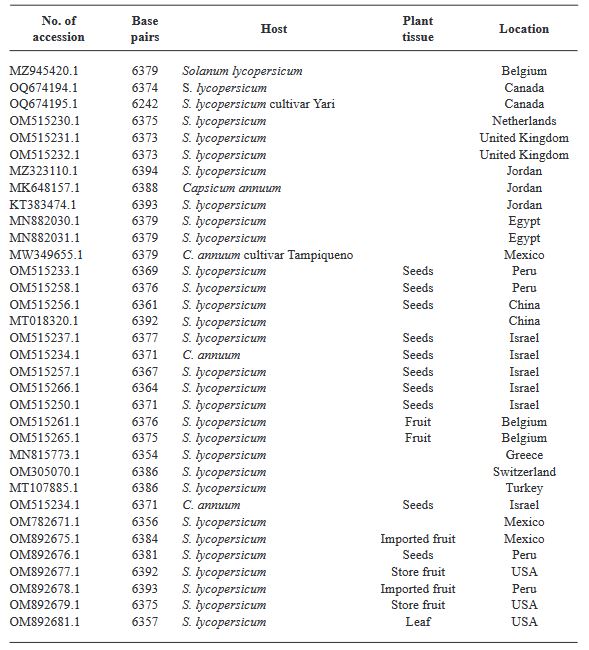
Materials and Methods. In greenhouses, the post-infection variability of two isolates was analyzed: EM-JI2021 (State of Mexico) and C-JI2021 (Colima) in 15 genotypes of tomato and 20 of pepper. Each isolate was mechanically inoculated on five plants per genotype with a total of 150 plants (56 days old) of tomato and 200 of pepper. Three plants per genotype were used as controls. Sixty-one days after inoculation, one leaf per plant was collected for RT-PCR. Incidence and symptom expression were recorded. RNA extraction was by 2% CTAB. ToBRFV-F/ ToBRFV-R primers amplifying 475 bpb of the RpRd gene were used (SENASICA-CNRF). 24 RT-PCR products were sequenced, cleaned and aligned with NCBI Genbank records using MEGAv11.0.13. Based on epidemiological criteria, 34 sequences were selected from GenBank for variability analysis.
Results. Ten days after inoculation, tomato genotypes exhibited severe mosaic, mild mosaic, and reduced leaf area. In pepper, symptoms differentiated by genotype were observed, including hypersensitivity reaction, leaf deformation, stem necrosis, mosaic, yellowing, necrotic lesions, and asymptomatic condition. Between position 2,124 to 2,500 bp there was 99.74 % homology with the first report of ToBRFV in Jordan (KT383474.1). Homology >99.74 % was found with isolates from USA (MT002973.1) and Canada (OQ674195.1). C-JI2021 exhibited no variability, while EM-JI2021 generated three haplotypes: One nucleotide change (c.2,355T>C) was detected in Mulato (pepper) and Don R (tomato), while two substitutions (c.2,278A>T; c.2,355T>C) were detected in Santawest, Altius, Sahariana and Nebula (tomato).
Conclusion. The pathogenic intensity of ToBRFV varied from asymptomatic to severe depending on the combination of host, genotype, and haplotype. In short periods of infection, three haplotypes were detected, suggesting host-dependent mutagenic capacity of the virus.
Share this link via
Or copy link


byEric Ángel Mendoza Pérez, Ricardo Santillán Mendoza, Humberto Estrella Maldonado, Cristian Matilde Hernández, Felipe Roberto Flores de la Rosa*, Jacel Adame García
Received: 18/July/2023 – Published: 06/March/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2307-6
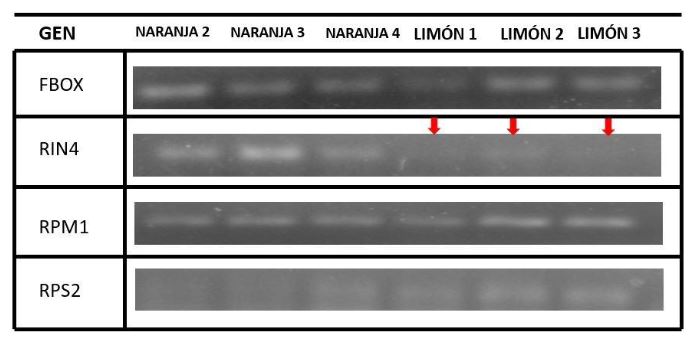
Abstract Objetive/Antecedents. Persian lime (Citrus latifolia) shows a very high level of tolerance to Huanglongbing (HLB). A recent study suggests that genes from the RPM1-RIN4-RPS2 complex could be partly responsible for HLB tolerance in Persian lime, unlike other highly susceptible species such as orange (C. sinensis). The objective of this study was to compare the expression of this gene complex between orange, highly susceptible to HLB, and Persian lime, a tolerant species.
Materials and Methods. Sequences of the three genes of the complex for orange and Persian lime were obtained from databases of previously published works, alignments and primer design for gene expression were performed using various bioinformatics tools. Subsequently, tissue samples from symptomatic HLB-infected orange and Persian lime were obtained and infection was confirmed. The expression of the RPM1-RIN4-RPS2 genes was compared using endpoint RT-PCR.
Results. The presence of all three genes of the complex was determined in both orange and Persian lime, and it was also determined that they are highly conserved between both species. Additionally, it was observed that there is no differential expression for the RPM1 gene in symptomatic HLB tissue; however, there is a difference in the expression of the RPS2 and RIN4 genes.
Conclusion. The results suggest that the contrasting response to HLB could be associated with the activity of the interaction of the RIN4 and RPS2 genes, thus, this could be of interest for citrus genetic improvement aiming at HLB control.
Share this link via
Or copy link

byCandelario Ortega Acosta, Reyna Isabel Rojas Martínez*, Daniel L. Ochoa Martínez, Manuel Silva Valenzuela
Received: 02/October/2023 – Published: 06/March/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-2
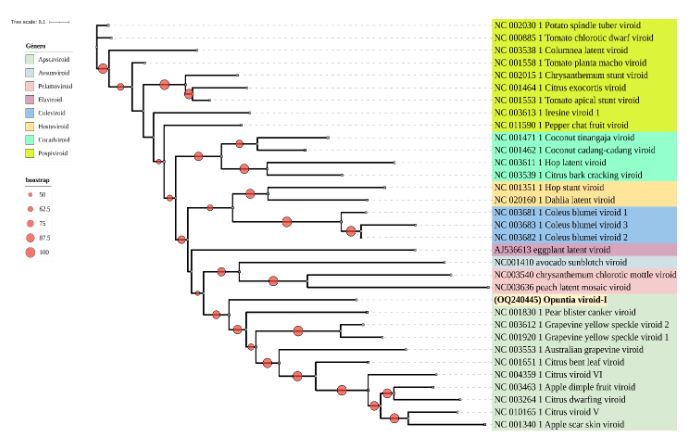
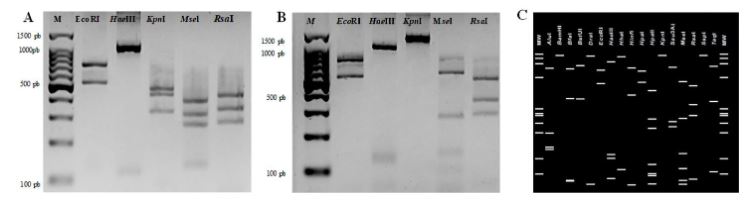
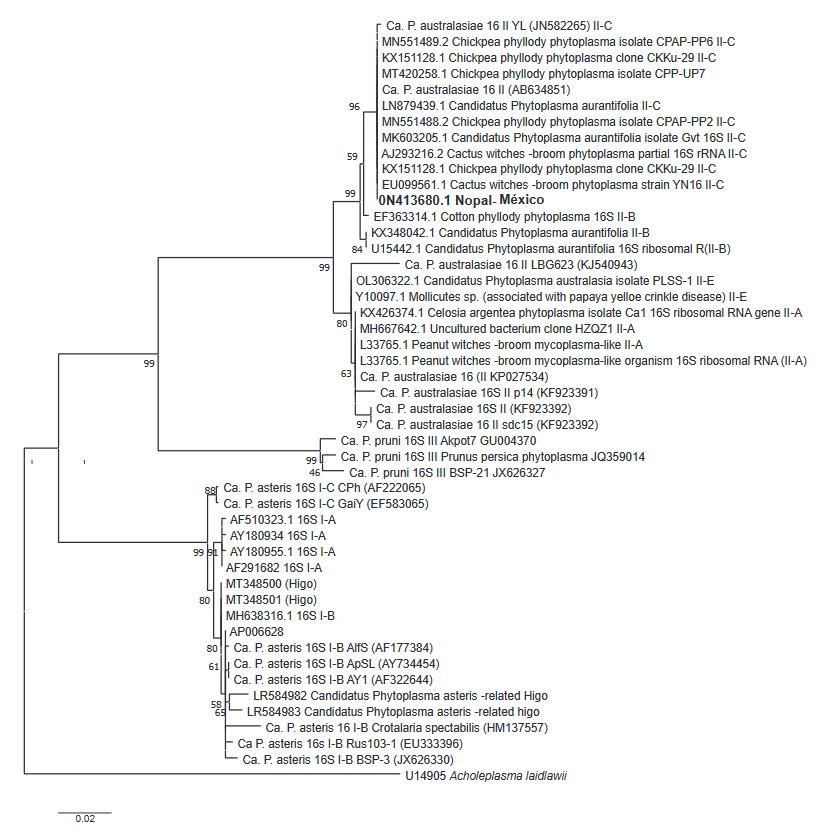
Abstract Background/Objective. Phytoplasmas are obligate plant pathogens that exhibit strong specificity with their hosts. Typical symptoms induced by these pathogens include stunted growth and general decline, among others, and they rarely lead to plant death. The aim of this research was to determine the phytoplasma associated with the ‘witch’s broom’ symptom in an ornamental cactus (Opuntia sp.).
Materials and Methods. Four samples of ornamental cacti exhibiting ‘witch’s broom’ symptoms were collected from four commercial nurseries in Texcoco, State of Mexico. DNA extraction was performed on the samples, followed by PCR using specific primers for phytoplasmas (P1/P7 and R16F2n/R16R2). Phytoplasma determination was carried out through PCR, in vitro RFLP, sequencing, and phylogenetic analysis.
Results. According to the various analyses conducted, it was determined that the phytoplasma associated with the ornamental cactus belongs to the subgroup 16SrII-C.
Conclusion. Based on the obtained results, it is established that a phytoplasma from the 16SrII-C subgroup is associated with the ‘witch’s broom’ symptom in the ornamental cactus (Opuntia sp.).
Share this link via
Or copy link

byLorena Uribe Lorío*, Lidieth Uribe, César Rodríguez, Luis Felipe Aráuz
Received: 26/May/2023 – Published: 26/February/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2305-5
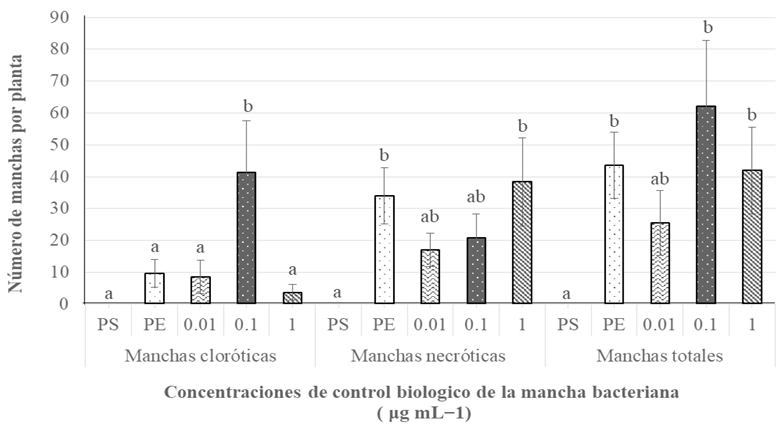
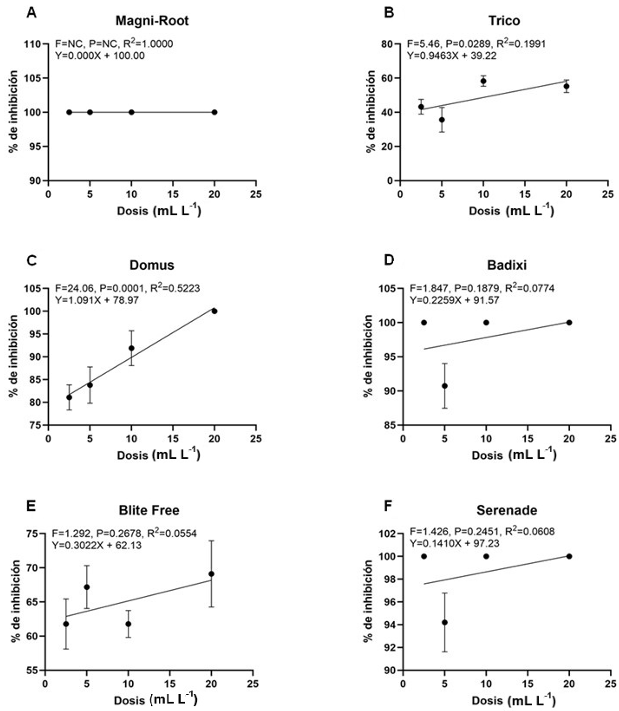
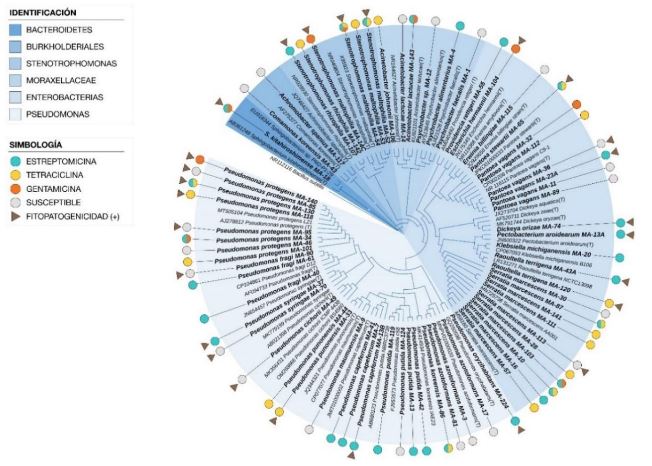
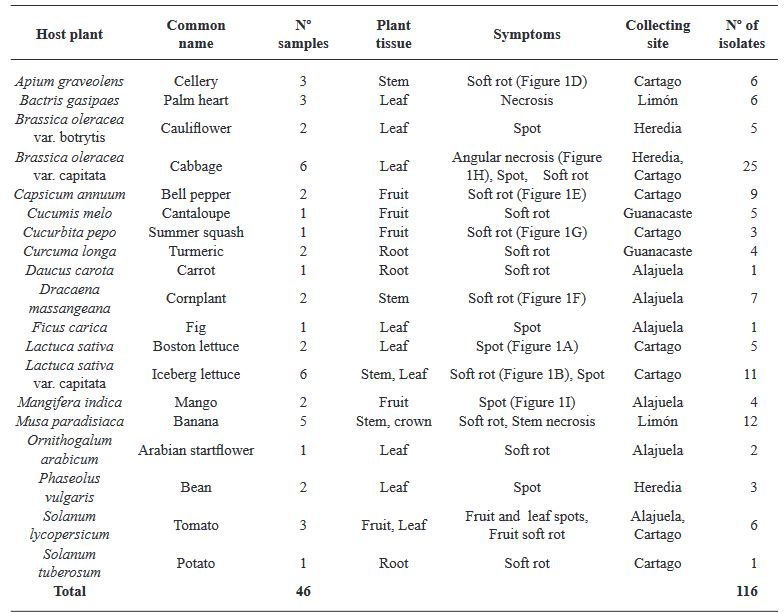
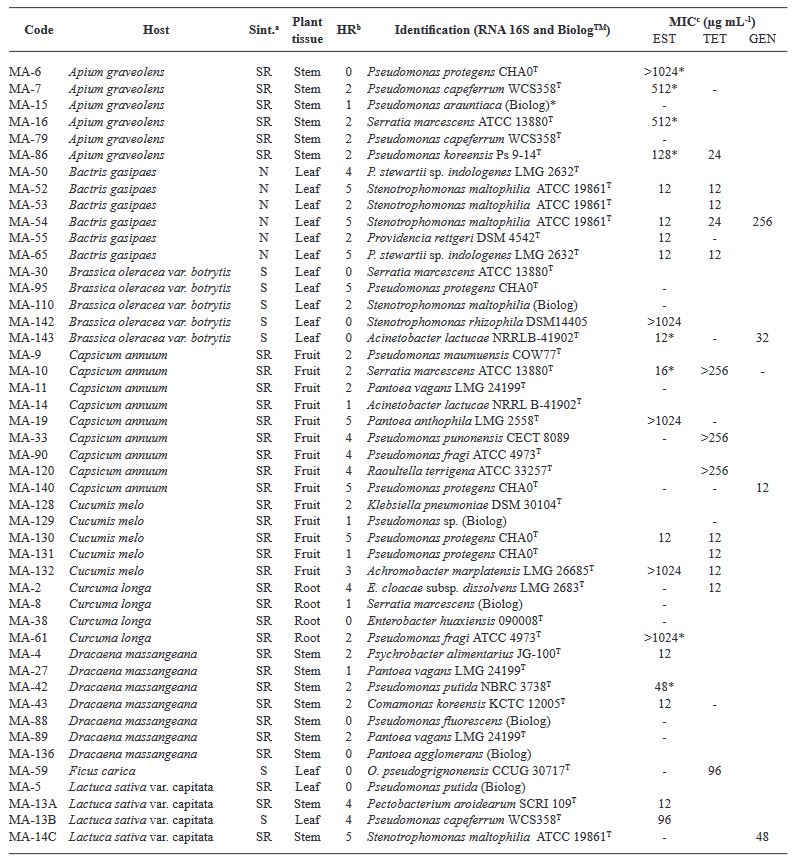
Abstract Objetive/Background. The aim of this was to assess the diversity and antibiotic resistance of bacteria isolated from 19 crops with bacterial infection symptoms.
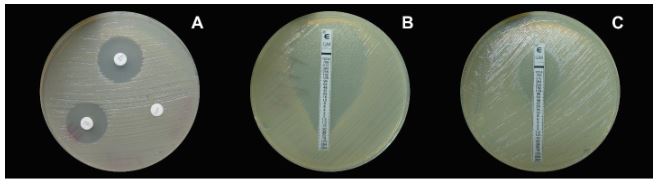
Materials and Methods. This collection was identified using 16S rRNA gene sequencing and the Biolog system. Susceptibility and minimum inhibitory concentration (MIC) for streptomycin, tetracycline, and gentamicin were determined using disk diffusion and E-test methods, respectively.
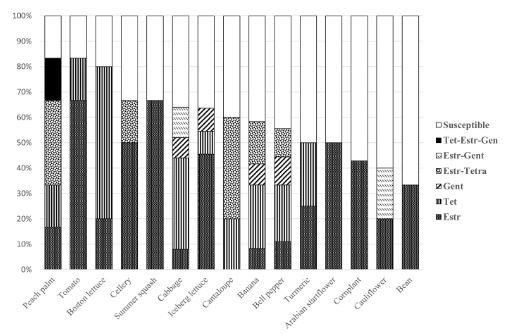
Results. A total of 55 species belonging to 20 bacterial genera were identified, with Pseudomonas, Serratia, Pantoea, and Stenotrophomonas being the most abundant. Approximately 27% of the isolates were categorized as pathogenic through the hypersensitivity reaction test, including phytopathogenic species like Pseudomonas syringae, P. cichorii, Pantoea anthophila, P. stewartii, Stenotrophomonas maltophilia, Dickeya oryzae, Erwinia billingiae, Pectobacterium aroidearum, and Enterobacter cloacae subsp. dissolvens. Resistance to at least one antibiotic was detected in 60% of isolates from 17 crops, with tomatoes, heart of palm, and lettuce exhibited the highest proportion of resistant bacteria (>80%). Streptomycin resistance was most common (35%), followed by tetracycline (28%) and gentamicin (9%).
Conclusion. The findings indicate the presence of antibiotic resistance in saprophytic and pathogenic bacteria associated with 17 out of 19 assessed crops, posing risks to the environment, phytosanitary conditions, and public health
Share this link via
Or copy link

byMaría José Armenta Rojas, Norma Ávila Alistac, María del Carmen Zúñiga Romano, Gerardo Acevedo Sánchez, Alfonso Muñoz Alcalá, Rene Gómez Mercado, Juan José Coria Contreras, Diana Gutiérrez Esquivel, Serafín Cruz Izquierdo, Ivonne García González, Oscar Bibiano Nava, Gustavo Mora Aguilera*
Received: 30/October/2023 – Published: 12/February/2024 – DOI: https://doi.org/10.18781/R.MEX.FIT.2310-7
Abstract Introduction/Objective. Ayocote bean (Phaseolus coccineus) has potential as a source of resistance in breeding programs because it exhibits greater tolerance to plant pathogens than P. vulgaris. However, its sanitary characterization is insipient; therefore, the purpose of this work was to carry out an etiological-epidemiological diagnosis, with emphasis on presumptive symptoms of viral and phytoplasmic organisms, and a typical fungal signs of powdery mildew.
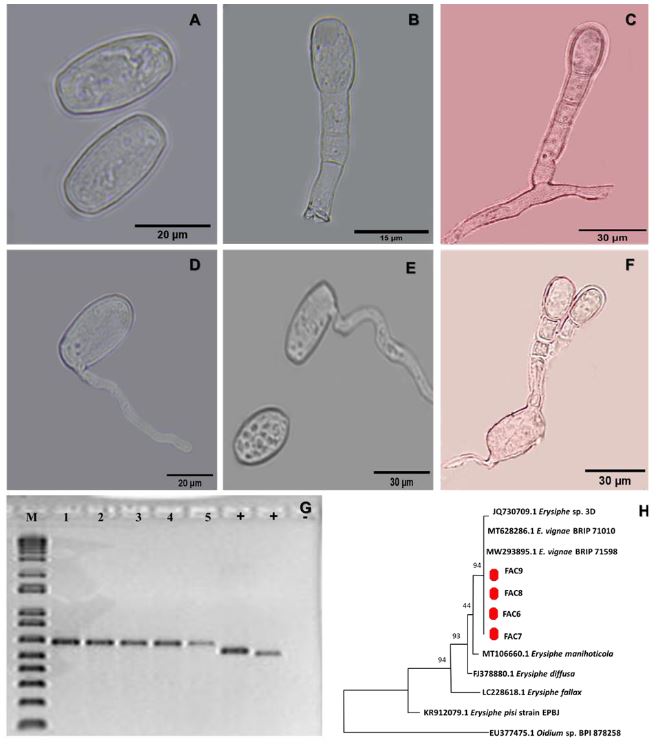
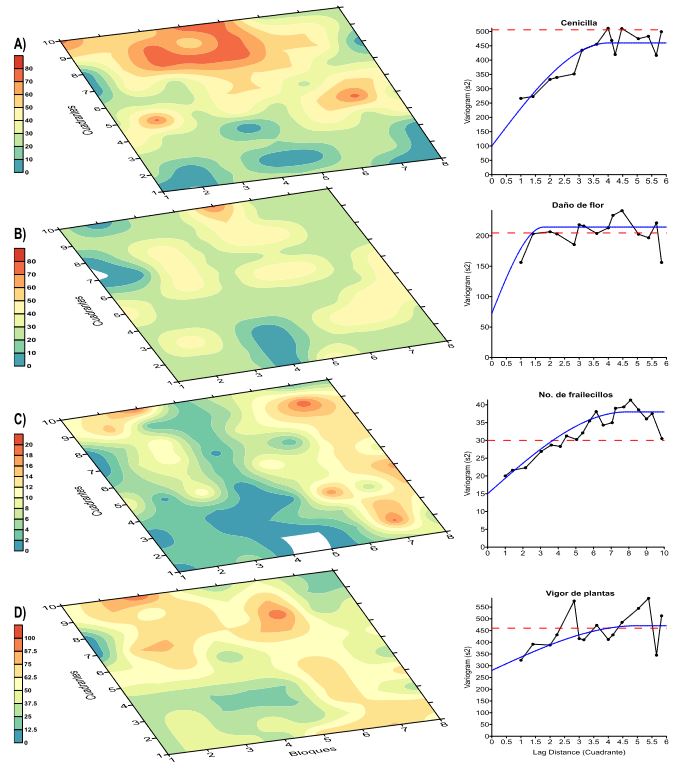
Materials and Methods. A plot (50 x 62 m) of flowering Ayocote bean was selected. It was divided into 80 (8 x 10) quadrats (6 x 6 m) and 720 subquadrats (2 x 2 m). From 25 plants with powdery-mildew-type leaf symptoms, mycelium was collected with adhesive tape for light microscopy observation and taxonomic identification. Length-width measurements were made on 60 conidia. Pure mycelium collected in situ and ex situ from 1-5 leaflets/plant was used for genomic analysis by PCR with universal primers ITS1 and ITS4. Samples were sequenced in Macrogen Inc. Korea. A total of 63 plants and 121 trifoliate leaves with viral and phytoplasmic symptoms were collected by direct sampling. In 88/121 samples, genomic analysis was performed by PCR with universal primers for Potyvirus (1), Begomovirus (2), and Phytoplasmas (1). Sequence editing and analysis were performed in SeqAssem and BLASTn/GenBank. Phylogenetic constructions were developed in Mega 11 with MUSCLE, Maximum Likelihood (ML), and HKY substitution model (1000-Bootstrap). Putative powdery mildew severity (%), flower damage (%), Macrodactylus sp. adult density, and plant vigor (%) were evaluated in 80 quadrats (3subquadrats/quadrat) with App-Monitor®v1.1 configured with a 5-class scale. In GoldenSurfer® v10, Kriging geostatistical analysis was performed to determine the spatial interrelationship between these variables.
Results. Erysiphe vignae was identified as associated with powdery mildew of P. coccineus. The fungus, with hyaline, ovoid to ellipsoid conidia measuring 31.74 ± 0.3419 μm x 15.11 ± 0.1579 μm, without the presence of fibrosin bodies, had 100% genomic homology. This is the first report in Mexico. With average July-August temperature and relative humidity of 16.3 °C (±5.8) and 92.8 % (±10.7), respectively, powdery mildew leaf incidence and severity were 65.3 and 22.7 % (±16.9, range: 0 - 66.5 %), respectively. The most inductive focus (60- 80 % severity) had an aggregate e 4-quadrat pattern (96 m2, lag = 4 and σ2-s = 450). Inoculum dispersal was significantly associated with dominant North-South winds and plant vigor (lag = 4 and σ2-s = 470). Flower damage was inconclusive in its spatial association with powdery mildew and Macrodactylus sp. suggesting uncorrelated events. No Potyvirus, Begomovirus, or Phytoplasmas were detected associated with yellowing, leaf distortion, mosaic, internode shortening, and other symptoms observed in situ. This confirms the relative tolerance/resistance reported for P. coccineus.
Conclusion. E. vignae (Erysiphales: Erysiphaceae) associated with P. coccineus is reported for the first time in Mexico with moderate to intense epidemic level, which indicates its susceptible condition to this fungus. However, negative results for Potyvirus, Begomovirus, and Phytoplasmas, validate the apparent tolerance/ resistance of P. coccineus to these organisms.
Share this link via
Or copy link


byMauricio Montero Astúa*, Izayana Sandoval Carvajal, Lisela Moreira Carmona, William Villalobos Muller, Laura Garita Salazar, Sofía Carvajal Rojas
Received: 30/July/2023 – Published: 30/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-3
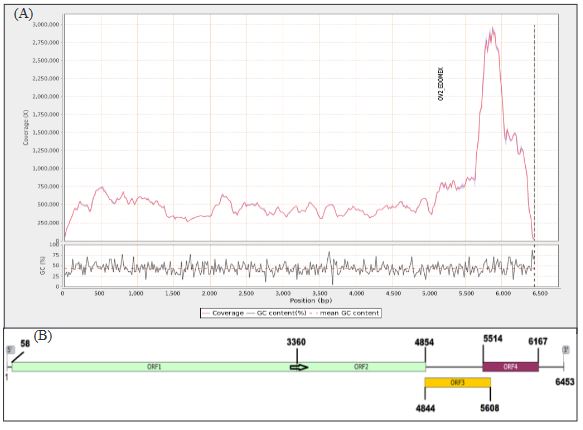
Abstract Background/Objective. Leaves of the shrub chaya (Cnidoscolus aconitifolius), spinach tree or ‘chicasquil’ (in Costa Rica), are consumed in the Mesoamerican culinary tradition, having its origin in South Mexico and Guatemala. The objective of this work was to verify the viral nature of the observed in a chaya plant disease and to identify the species of the virus.
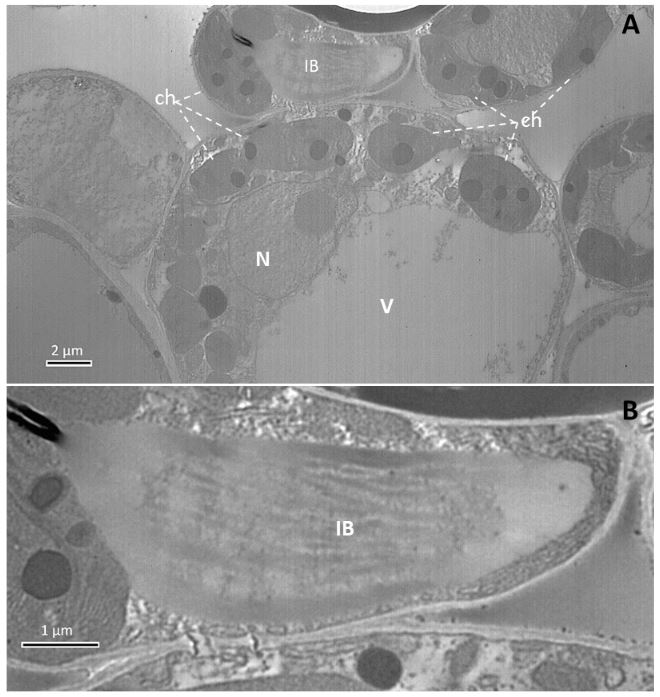
Materials and Methods. Plant virus detection and identification was achieved by TEM, RT-PCR using degenerated primers to potexviruses, and sequencing. Pathogenicity tests were done by mechanical inoculation using chaya symptomatic tissue, on Nicotiana benthamiana and chaya plants.
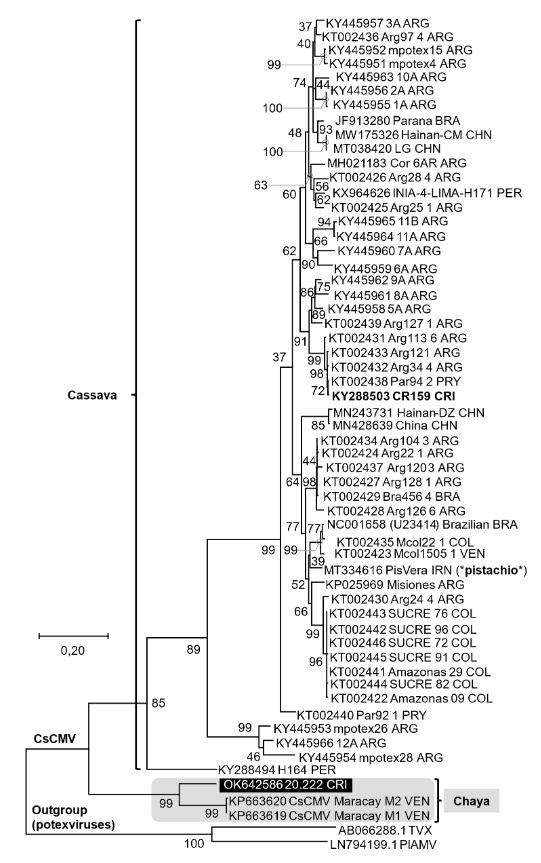
Results. We report CsCMV detection in a chaya plant in Costa Rica with mosaic symptoms. Pathogenicity and association of virus and symptoms were demonstrated by mechanical inoculation in Nicotiana benthamiana and chaya plants. We hypothesize this infection corresponds to a recent introduction and discussed how cultural traditions impact the distribution of plant viruses.
Conclusion. The findings confirm the presence of a CsCMV-related virus, previously unreported for Costa Rica, in Cnidoscolus aconitifolius. The results herein highlighted the need to study its distribution and diversity throughout Latin America
Share this link via
Or copy link

Comparison of molecular protocols to detect Tomato brown rugose fruit virus in solanaceae hosts
byErika J. Zamora Macorra, Katia Aviña Padilla*, Rosemarie W Hammond, Daniel L. Ochoa Martínez
Received: 23/August/2023 – Published: 23/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-5
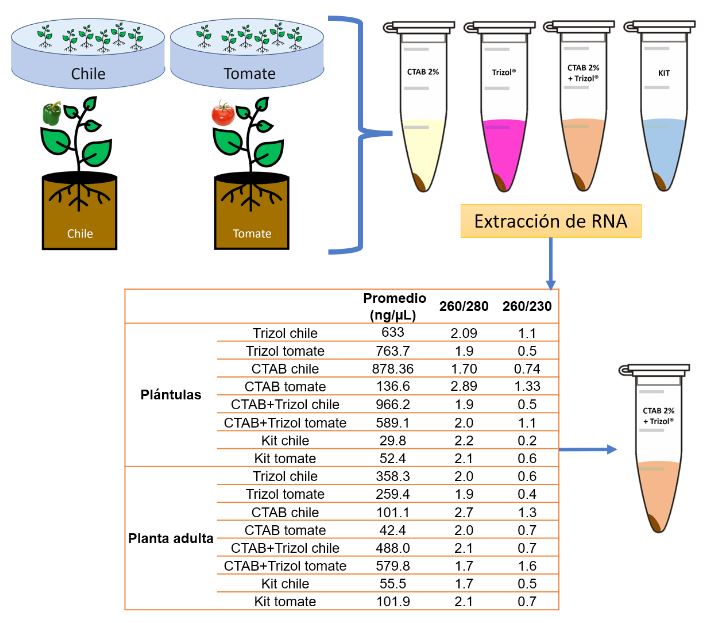
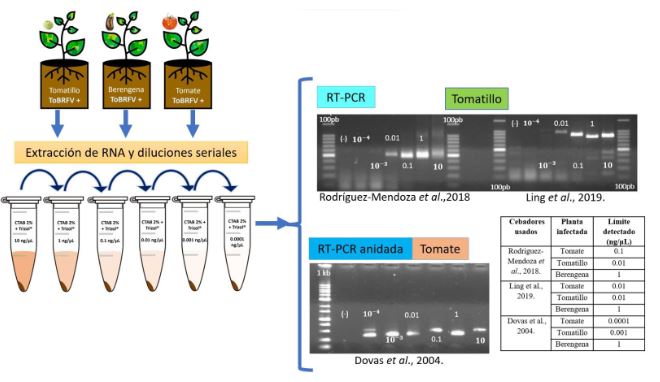
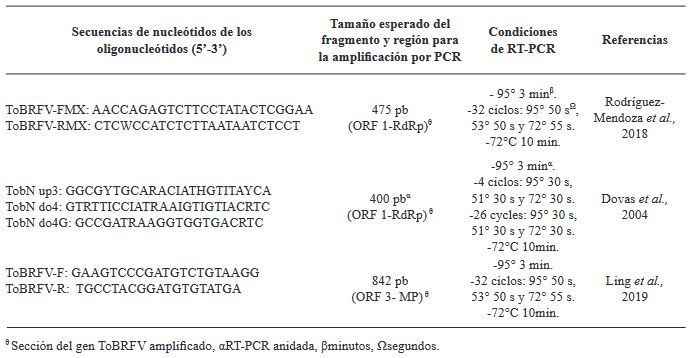
Abstract Background/Objective. The Tomato brown rugose fruit virus (ToBRFV) has emerged as a significant threat to Solanaceae crops, including tomato and pepper. Its presence in Mexico since 2018 has raised concerns about its impact on agricultural production. Early and accurate detection of this pathogen is essential to prevent its spread and mitigate its effects. In Mexico, several molecular techniques are employed for its diagnosis, including endpoint RT-PCR, RT-qPCR, and multiplex RT-qPCR.
Materials and Methods. This research aimed to assess the efficiency of different RNA extraction methods in combination with specific PCR primers for detecting ToBRFV.
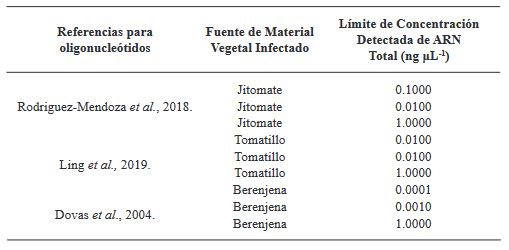
Results. Among the methods tested, the CTAB-Trizol RNA extraction protocol combined with nested PCR using primers reported by Dovas et al. (2004) was identified as the most sensitive molecular method for detecting the virus.
Conclusion. This finding highlights the importance of selecting the appropriate combination of extraction and amplification protocols to achieve optimal sensitivity and accuracy in ToBRFV detection.
Share this link via
Or copy link

byCarlos D. Ramos Villanueva, Guadalupe Carrillo Benitez, Erika J. Zamora Macorra*, Eduardo Santiago Elena, Samuel Ramírez Alarcón, Jezrael Jimenez Vidals, Ricardo Ricardo López
Received: 31/July/2023 – Published: 19/December/2023 – DOI: https://doi.org/10.18781/R.MEX.FIT.2023-1
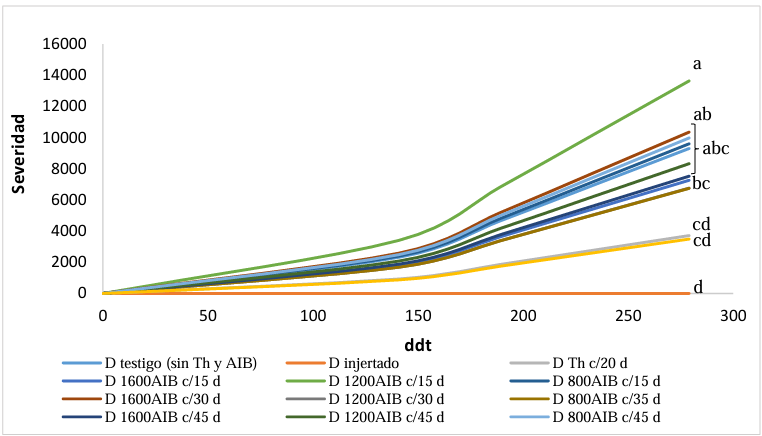
Abstract Background and objective: The Tomato brown rugose fruit virus (ToBRFV) is one of the main pathogens affecting tomato crops in Mexico. Despite efforts to prevent its spread, it is nearly impossible due to its low transmission percentage through seeds and its high susceptibility to being transmitted through cultural practices. Therefore, alternative management strategies are being sought. This research aimed to determine the effect of endophytic microorganisms applied to the soil on tomato plants infected with ToBRFV.
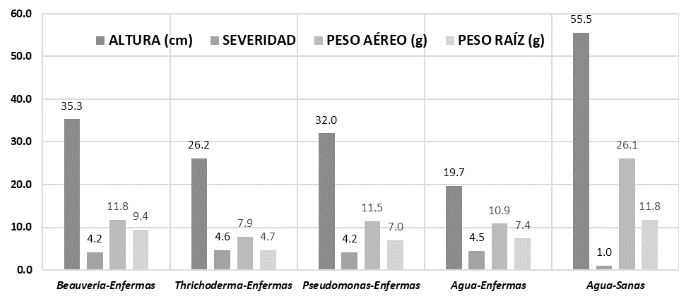
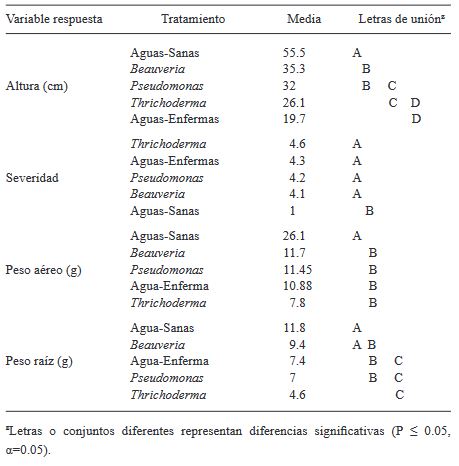
Materials and Methods. A tomato plant was used as an experimental unit, with 13 repetitions per treatment. The treatments on tomato plants infected with ToBRFV were Beauveria peruviencis, Trichoderma longibrachiatum, Pseudomonas sp. and water as a sick witness; a treatment of healthy plants treated with water was also included as an absolute control. The response variables were plant height, fresh weight of the aerial part and root and severity (two evaluations). Measurements were analyzed using Tukey-Kramer HSD tests for each pair. Results and conclusion: Significant differences were found Beauveria peruviencis, Trichoderma longibrachiatum, Pseudomonas sp. and water as a sick witness. The treatment that most favored the development of infected plants (79% taller and 15% heavier than infected mock) and reduced its severity was B. peruviensis, followed by Pseudomonas sp. On the other hand, the treatment that resulted in the least plant development (31% smaller than infected mock) and even increased the severity of the infection was T. longibrachiatum.
Share this link via
Or copy link